
Cómo citar: Alvarado-Ortega, J., Porras-Múzquiz, H.G., 2022, On the presence of Xiphactinus (Teleostei, Ichthyodectiformes) in the Coniacian-Campanian marine deposits of the Piedritas site, Coahuila, northern Mexico: Revista Mexicana de Ciencias Geológicas, v. 39, núm. 2, p. 142-150.
REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS
v. 39, núm. 2, 2022, 142-150
DOI: http://dx.doi.org/10.22201/cgeo.20072902e.2022.2.1684
On the presence of Xiphactinus (Teleostei, Ichthyodectiformes) in the Coniacian-Campanian marine deposits of the Piedritas site, Coahuila, northern Mexico
Jesús Alvarado-Ortega1,* and Héctor Gerardo Porras-Múzquiz2,**
1 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Col. Copilco, Del. Coyoacán, Ciudad de México, 04510 Mexico.
2 Museo de Paleontología de Múzquiz, Adolfo E. Romo 1701, La Cascada, 26343, Santa Rosa de Múzquiz, Melchor Múzquiz, Coahuila, Mexico.
ABSTRACT
The discovery of the first Mexican fossil specimen belonging to the species Xiphactinus audax is reported here. This specimen was recovered in the Coniacian-Campanian marine deposits of the Austin Group at the Piedritas site, within the homonymous ejido, Municipality of Ocampo, north of Coahuila. Although the specimen is strongly fragmentary, partially preserved by impressions or molds of the bones, and only shows parts of the head, the opercular series, and the pectoral fins, this has enough anatomical features that allow its specific taxonomic determination. Among these diagnostic features, this fossil has a well-developed ethmopalatine bone, a well-developed supraoccipital crest, a maxilla anteriorly higher than the rest of the bone, and a very robust palatine articular head. Particularly, this fossil exhibits two distinctive features of the genus Xiphactinus, a row of conical teeth, irregularly sized and distributed in the maxilla and dentary bones, as well as a pectoral radius 1 extremely developed and at least 2.5 times wider than the subsequent ray. At the same time, its teeth are smooth and have no sharp edges or carenae, which allows its specific identification. The present discovery complements the geographical distribution of Xiphactinus confirming its wide longitudinal distribution throughout America.
Key words: Xiphactinus; Ichthyodectiformes; Late Cretaceous; Piedritas; Coahuila; Mexico.
RESUMEN
Se reporta el hallazgo del primer ejemplar fósil mexicano perteneciente de la especie Xiphactinus audax, recuperado en los depósitos marinos del Coniaciano-Campaniano del Grupo Austin del sitio Piedritas, dentro del ejido homónimo, Municipio de Ocampo, al norte Coahuila. A pesar de que el ejemplar es fragmentario, en partes conservado por impresiones o moldes de los huesos y solo conserva partes de la cabeza, la serie opercular y la cintura y aletas pectorales, en este se conservan suficientes rasgos anatómicos que permiten su determinación taxonómica especifica. Entre estos rasgos diagnósticos de este fósil es posible observar la presencia de un hueso etmopalatino bien desarrollado, una cresta supraoccipital grande, un maxilar anteriormente más alto que el resto del hueso y una cabeza articular del palatino muy robusta. De manera particular, este fósil exhibe dos rasgos distintivos del género Xiphactinus, una hilera de dientes cónicos, de talla irregular e irregularmente dispersos en la maxila y mandíbula, así como un radio pectoral 1 peculiarmente muy desarrollado y al menos 2.5 veces más ancho que el siguiente radio de esta aleta. Al mismo tiempo, los dientes de este pez son lisos y no presentan bordes cortantes o carenas, lo que permite su determinación especifica. Este hallazgo complementa la distribución geográfica de Xiphactinus y confirma su amplia distribución longitudinal a través de toda América.
Palabras clave: Xiphactinus; Ichthyodectiformes; Cretácico Tardío; Piedritas; Coahuila; México.
Manuscript received: february 24, 2022
Corrected manuscript received: april 22, 2022
Manuscript accepted: april 23, 2022
INTRODUCTION
Due to its huge size and fierce appearance, Xiphactinus Leidy, 1870, is the most outstanding fish of the order Ichthyodectiformes. This Bathonian-Maastrichtian clade erected by Bardack and Sprinkle (1969) includes extinct marine teleosts that today involve about 32 genera. Two synapomorphies support the monophyly of the clade, the presence of the ethmopalatines forming the floor of the nasal capsule plus the uroneurals covering the lateral surface of the last preural and both ural centra (Patterson and Rosen, 1977). The bauplan of these fishes also includes the bulldog aspect of the head; the moderate to very elongated trunk; the wide-saber shape of the first rays in the paired fins; the large and expanded coracoids having a longitudinal symphysis; the paired fins opposed and located far back in the body; and a comparatively short dorsal fin.
During the Jurassic, the ichthyodectiformes emerged as small predators with relative short bodies and delicate jaws; however, since the Early Cretaceous their bodies became longer at the same time as their food habits diversified thanks to the development of stronger jaws and the diversification of the teeth shapes. In the Late Cretaceous, some ichthyodectiforms became top predators, such as Xiphactinus, which possessed the largest and strongest teeth of the order and its length reached about six meters, turning it into the largest teleost of that time. The ichthyodectiforms disappeared in the Maastrichtian, near the Cretaceous–Paleogene mass extinction event (Bardack, 1965; Schaeffer and Patterson, 1984). Fossil remains of these fishes have been collected in transitional to open marine deposits of the epicontinental seas around the world, in America, Europe, Africa, Asia, Australia, and Antarctica (Patterson and Rosen, 1977; Arratia et al., 2004; Berrell et al., 2014; Kim et al., 2014; Yabumoto et al., 2018; Bahrami et al., 2021).
In Mexico, the fossil record and diversity of the ichthyodectiforms have increased in the last twenty years. Here, six nominal species plus other indeterminate specimens belonging to six genera have been already identified, and the studies of many specimens are in progress. Such taxonomical diversity reveals this country as a hotspot to understand the natural history of the Ichthyodectiformes (Alvarado-Ortega, 1998, 2005). Prymnetes longiventer Cope, 1871, is a mysterious Cretaceous species known by a single specimen from an undocumented site, near Tuxtla Gutierrez, Chiapas. Felix (1891) reported the presence of Thrissops Agassiz, 1833, near Tlaxiaco, Oaxaca, based on small and fragmented bones that today have no diagnostic value; however, this fish seems present in the Kimmeridgian deposits of Mazapil, Zacatecas (Villaseñor et al., 2006). Maldonado-Koerdell (1956) reported the presence of Xiphactinus and Ichthyodectes Cope, 1870, in the Turonian sequence of the Agua Nueva Formation exploited in the Xilitla quarry; unfortunately, his fossils are missing, were observed on rocks out of geological context, and the anatomical parts described then have no diagnostic features. The most important paleontological site of the Agua Nueva Formation is the Vallecillo quarry, Nuevo León, where complete specimens of Gillicus arcuatus (Cope, 1875), Vallecillichthys multivertebratum Blanco and Cavin, 2003, and an indeterminate species of Heckelichthys Taverne, 2008, have been recovered (Blanco-Piñón and Alvarado-Ortega, 2007; Giersch, 2014; Baños-Rodríguez, et al, 2020). Unamichthys espinosai Alvarado-Ortega, 2004, so far is the only species described from the Albian deposits of the Tlayúa Quarry, Puebla. Specimens of Gillicus arcuatus and Saurodon leanus Hays, 1830, also have been collected in the Eagle Ford Formation at the La Mula, Los Pilotes Ranch, and other Turonian-Coniacian sites near Múzquiz, Coahuila (Alvarado-Ortega et al., 2006a; Alvarado-Ortega and Porras-Múzquiz, 2009; Giersch et al., 2008; Giersch, 2014). Saurodon Hays, 1830, also is present in the Campanian deposits of the Tzimol quarry, near Ochuxhob, Chiapas (Alvarado-Ortega et al., 2020). Heckelichthys opercularis Baños-Rodríguez, González-Rodríguez, Wilson, and González-Martínez, 2020, and fragmented specimens that probably belong to Unamichthys Alvarado-Ortega, 2004, are present in the Albian deposits belonging to the El Doctor Formation exploited in the Muhi quarry, Hidalgo (Alvarado-Ortega, 2005; Alvarado-Ortega et al., 2006c; Baños Rodríguez, 2018). Numerous remains of still indeterminate ichthyodectiforms have been collected in Xilitla and Tlayúa quarries, as well as in the Early Cenomanian deposits of the El Chango quarry (Cintalapa Fo......rmation, Chiapas) and the Turoninan Arroyo las Bocas site (Mexcala Formation, Guerrero) (Alvarado-Ortega, 2005; Alvarado-Ortega et al., 2006a, 2006b, 2006c, 2009; Than-Marchese and Alvarado-Ortega, 2011).
Although Alvarado-Ortega (2005) denied the presence of Xiphactinus in Mexico based on the insufficient evidence provided by Maldonado-Koerdell (1956); recently, Alvarado-Ortega and Porras-Múzquiz (2015) reported the first indisputable remains of Xiphactinus in this country. Such a rediscovery of Xiphactinus in Mexico is based on the fragments of the head and pectoral fin belonging to a large, incomplete, and single specimen recovered in the Coniacian-Campanian deposits of Austin Group exploited in the Piedritas site, Coahuila. This paper aims to provide an accurate description of this large specimen and explore its taxonomical and biogeographical implications.
GEOLOGICAL SETTINGS
The Piedritas site is an area of about 3 km2 within the Ejido Piedritas, Ocampo Municipality, Coahuila State, northern Mexico, about 20 km south of the Mexico-USA border (Porras-Múzquiz et al., 2014). In this site located between 28°48’15.5” N–103°6’5” W, at 769 m a.s.l., slabs of limestone are eventually extracted from small quarries opened by local people in the low hills along the east side of Boquillas Creek (Figures 1, 2).
In Piedritas, the fossiliferous sequence consists of about 10 m thick of thin laminated yellowish-gray limestone, marl, and shale rhythmically interbedded belonging to the Austin Group. The marly limestone of this sequence alternates with centimeter-thick clay beds that bear iron hydroxides forming small lenses, nodules, and framboidal pyrite. Local workers have commercially exploited these strata since these are used to cover floors and walls. The fossil assemblage of this site includes well-preserved tests of planktonic foraminifers, and bivalves (including Inoceramus Sowerby, 1814, and Platyceramus Heinz, 1932), gastropods, ammonites, crustaceans, marine reptiles (mosasaurs and pterosaurs), and Ichthyornis Marsh, 1872. The fishes recovered at this site include unidentified chondrichthyan teeth, remains attributable to Ichthyodectiformes, clupeomorphs, and those already identified as Enchodus Agassiz, 1835; Pachyrhizodus Dixon, 1850; and Laminospondylus transversus Springer, 1957 (Riquelme et al., 2013; Porras-Múzquiz et al., 2014). The lithologic sequence and fossil assemblage of Piedritas are similar to those reported at other nearby sites, previously reported as the ‘Múzquiz Lagerstätte’ of Coahuila, in which deposits belonging to the Eagle Ford and Austin groups are involved (Alvarado-Ortega and Porras-Múzquiz, 2009, Blanco-Piñón and Alvarado-Ortega, 2005, Giersch et al., 2008, Stinnesbeck et al., 2005, Riquelme et al., 2013). Although the fauna at Piedritas has yet to be studied thoroughly, Porras-Múzquiz et al. (2014) concluded that this is part of the Coniacian-Early Campanian deposits of the Austin Group.
Figure 1. Map of Coahuila showing the position of the paleontological site of Piedrital in Coahuila, northern Mexico (modified from Riquelme et al., 2013, fig. 1).

Figure 2. A general view of the sedimentological sequence of the Austin Group exposed in the Piedritas site, northern Coahuila, Mexico.
MATERIALS AND METHODS
The specimen described here was prepared mechanically with air scribes of different potency and point diameters. Dust and dirt from the fossil and the rocky matrix surfaces were washed by immersion into a weak aqueous solution of Potassium Hydroxide for 30 min. Finally, this was washed into clean freshwater for 1 hr. and dried up in the shade. The weak and broken fragments of the fossil were glued with cyanoacrylate and hardened with a weak solution of plexigum and methacrylate acetate applied with fine brushes. The fossil was observed and photographed under different illumination conditions, under daylight, with white light before and after being coated with ammonium citrate, and long-wave UV (254 nm) light, as well as under dry and wet conditions.
Anatomical nomenclature
The anatomical nomenclatures and abbreviations used in this work follow those used in previous similar studies (e.g. Bardack,1965; Patterson and Rosen, 1977; Maisey, 1991).
Institutional abbreviations
The specimens referred to in the present work are housed in the following institutions: AMNH, American Museum of Natural History, New York. USA. IGM, Colección Nacional de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México. MUZ, Museo de Paleontología de Múzquiz, Coahuila, Mexico. UERJ-PMB, Paleontological Collection of the Universidad Estatual do Estado do Rio de Janeiro, by Dr. Paulo M. Brito. USNM, United States National Museum, Smithsonian Institute, Washington D.C., USA.
Comparative materials
The following specimens were included in the present research for comparison purposes. Chiromystus alagoensis Jordan, 1910: AMNH 10015 and AMNH 10016, both from Riacho Doce, Alagoas, Brazil. Cladocyclus gardneri Agassiz, 1841: AMNH 11877, AMNH 19129, AMNH 11992, AMNH 19528, UERJ-PMB 510, and UERJ-PMB 520; all from the Santana Formation, Ceará, Brazil. Cladocyclus sp.: AMNH 2982, AMNH 3875, AMNH, AMNH 12709,19166, and AMNH 19167, all from the Santana Formation, Ceará, Brazil. Gillicus arcuatus (Cope, 1875): AMNH 8571 from Niobrara, Kansas, USA; and MUZ 47a and MUZ 47b, from the El Pilote Ranch, Coahuila, Mexico. Ogunichthys triangularis Alvarado-Ortega and Brito, 2010: Type series including UERJ-PMB 100 (holotype), UERJ-PMB 93 to UERJ-PMB 99 plus DNPM 533-P, 534-P, and 544-P; all from the Marizal Formation, Bahía, Brazil. Prymnetes longiventer Cope, 1871: USNM 4090 (silicone peel deposited in IGM), from an unknown locality, Chiapas, Mexico. Proportheus kameruni Jaekel. 1909: AMNH 8394 and AMNH 6302, both from the San Benito River, Equatorial Guinea. Saurodon sp: IGM 6762, from the La Mula quarry, Mexico. Xiphactinus audax Leidy, 1870: AMNH 1673, AMNH 7350, AMNH 8547 AMNH 19528, all from Niobrara, Kansas, USA. Unamichthys espinosai Alvarado-Ortega, 2004: type series including IGM 8373 to IGM 8376, all from the Tlayúa quarry, Mexico.
RESULTS
Systematic Paleontology
Division Teleostei Müller, 1845.
Order Ichthyodectiformes Bardack and Sprinkle, 1969.
Suborder Ichthyodectoidei Romer, 1966.
Genus Xiphactinus Leidy 1870
Referred material. A single specimen, MUZ-3912, which preserves a fragment of the head and pectoral girdle exposing the left side, as well as the cross-section of the mandibular bones embedded in the rock and exposed in the thickness of the carrier rock (Figure 3).
Locality and age horizon. The Piedritas site, Ejido Piedritas, Coahuila, northern Mexico; upper Coniacian to lower Campanian Austin Group, (Porras-Múzquiz et al., 2014).
Description
General proportion. Although the head of MUZ-3912 is incomplete because the skull is dissociated from the opercular series; the head can be estimated at least 502 mm in length, including at least 312 mm of the length of the skull and 190 mm corresponding to the length of the opercle on its horizontal axis. Bardack (1965: p. 37) stated that in Xiphactunus the standard length contains 5.5–6 times its head length; this means that the standard length of this Mexican specimen was between 2761 and 3012 mm.
Skull. In MUZ-3912, the roof of the skull is almost fully exposed. In its most anterior end, the rostrodermethmoid is a robust structure, the anterior half of which is occupied by a rounded anterior process and a couple of prominent and rounded lateral processes that are project a little forward enclosing the anterior part of the nasal capsule. The posterior half of this bone widens a little caudally, does not show any longitudinal division, and is overlapped by the pair of frontal bones (Figure 3).
In addition to the rostrodermethmoid, the nasal skull region of MUZ-3912 shows the ethmopalatine and the lateroethmoid forming the floor and posterior part of the nasal capsule. Here, the lateroethmoid is a robust rectangular bone, widened at its dorsal and ventral ends and slightly bifurcated at the base. The ethmopalatine is slightly displaced from its life position; this elongated and robust bone exhibits part of its sinuous ventral surface, in which there are two wide and shallow articular facets, the anterior one articulates the anterior facet of the maxilla while the wider and posterior one corresponds the articular head of the palatine. Part of the vomer is observed below the left maxilla, ethmopalatine, and the articular head of the palatine, in the ventral position to the nasal capsule (Figure 3).
Of both frontals, only the left one is fully exposed. These bones occupy about one-third of the length of the skull. Anteriorly, these cover the posterior margin of the rostrodermethmoid. Laterally, these are somewhat widened and cover the orbit. And caudally, these extend to suture with the mesoparietal, pterotic, and sphenotic. Both frontals are entirely sutured medially through a weakly sinuous suture. The frontal surface is smooth except for a series of straight and shallow grooves that radiate everywhere from the bone center (Figure 3).
Although only the left part of the middle region of the skull roof is exposed, it is possible to recognize that this is occupied by two unpaired bones, the mesoparietal and supraoccipital, plus the pair of epioccipitals while its lateral region is occupied by the dorsal portion of the of pterotics. Here, the mesoparietal is a compound bone, resulting from the fusion of two parietal bones. The mesoparietal is narrow medially and deeply bifurcated posteriorly; it has a sharp anterior projection separating the posterior ends of both fronts a little; its posterior margin is concave and joins the anterior medial end of the supraoccipital; and each of its lateral extensions joins the respective epioccipital, suturing with about a quarter of the base of the supraoccipital and part of the dorsal edge of the pterotic. Medially, the mesoparietal elevates posteriorly forming the anterior part of the supraoccipital crest (Figure 3).
The supraoccipital bone is long and narrow. Dorsally, this forms the largest part of a well-developed supraoccipital crest; this triangular, broad, and laminar structure shows a convex anterior edge (and probably, its posterior was concave). Although this crest is incomplete in MUZ-3912; it is as high as the rest of the postorbital region of the skull and occupies at least the posterior half of the skull roof, from the posterior level of the orbit, and extends further back than the base of the skull (Figure 3).
The epioccipital is an elongated bone that anteriorly is sutured to the mesoparietal and posteriorly placed between the lateral edge of the supraoccipital and the mediodorsal edge of the pterotic. The fragment of a large laminar bone, here interpreted as the left posttemporal, covers the left side of the posterior end of the skull, including the posterior region of the supraoccipital crest and the posterior part of the left epioccipital (Figure 3).
In the temporal region of the skull, this specimen only exhibits the sphenotic, pterotic, and part of the intercalar. The sphenotic is a stout massive bone that borders the dorsoposterior part of the orbit and bears a stout lateral process projected outward at the level of the middle orbit. The pterotic is the largest bone in this part of the skull, this shows a long groove forming the dilator fossa, which encloses its suture with the sphenotic. A small part of the intercalary sutures the posterior end of the pterotic (Figure 3).
Circumorbital bones. In MUZ-3912 preserves a few remains of those bones of the circumorbital ring. Part of the large left infraorbital (proba-bly, the infraorbital 3) is preserved covering a large part of the cheek; the edges of this laminar bone are missing but its surface is peculiarly irregular and covered by small and round dome-like folds like those of the preopercle. Also, part of a right infraorbital (probably, the infraorbital 2) is present but separated from the head. The dermosphenotic bone is a rectangular flat bone that borders the dorsoposterior corner of the orbit. The supraorbital bone is a flat and rectangular structure that borders the anterior dorsal half of the orbit. At the center of the orbit, there is a uniformly wide and curved bone, which by its position corresponds to the anterior one of the sclerotic bones. No remains of the basal sclerotic bone are observed (Figure 3).
Jaw. The bones preserved in the jaw of MUZ-3912 include fragments and molds of teeth of maxillae and mandibular bones. The dorsal edge and large part of the anterior part of the left maxilla are preserved, revealing that this was a wide and stout flat structure with two anterior dorsal processes and a posterior straight dorsal edge. The anterior region of the maxilla carries both articular processes and is higher than the posterior part of this bone and shows a shallow depression in which the corresponding premaxilla was allocated; among these processes, the anterior is slightly shorter and in life could have been articulated the tip of the lateral process of the rostrodermethmoid and the anterior facet of the ethmopalatine; on the contrary, the posterior process is articulated with the ventral facet of the articular head of the palatine. This specimen exhibits the anterior of the right maxilla, where its anterior articular process rises. The alveolar edge of the left maxilla is poorly preserved and almost totally missing; however, it exhibits four alveoli of irregular size, and irregularly spaced, forming a single row. Two of these alveoli preserved the tooth root while the other two were unoccupied. The anterior tip of the right maxilla shows a small tooth conical, smooth, and slightly curved backward. The most anterior of these alveoli are placed below the region of the articular processes of this bone, which shows that the entire ventral region of the maxilla was toothed. No remains of any supramaxillae are preserved (Figure 3).
In MUZ-3912, both lower jaw branches are almost entirely missing; however, the ventral view of this specimen, shows that these preserve a few osseous remains and molds of parts of both dental bones and some of their respective teeth. In this view, the dentary bones are straight and thin structures, in which the anterior tips are slightly curved medially to form the dentary symphysis. As in the left maxilla, the alveolar edges of both dentary bones exhibit alveoli and tooth molds forming a single row. Such tooth cavities are circular to ovoid in shape, irregular sized, and unevenly spaced. In the left dentary, the transversal sections of two teeth are preserved showing a rounded, smooth outer surface and have no evidence of any carenae or ornamentation. The difference in the shape of these cavities is interpreted here as an artifact of the differential compression experienced by the specimen during its burial, which could not affect the alveoli size revealing that this fish had teeth of very different sizes (Figure 3).
Suspensorium. Most bones of this bone series are lots in MUZ-3912. Anteriorly, the palatine bone is recognized by its robust articular head that is rectangular, high, and shows the facet for the ethmopalatine dorsally and posterior articular process of the maxilla ventrally. Possible fragments of the ectopterygoid and mesetmoid bones are present between the orbit and left maxilla (Figure 3).
Opercular series and Brachostegal Rays. In MUZ-3912, the preopercle and infraopercle are not preserved; however, the left opercle and subopercle are well-preserved. The opercle is a flat and kidney-shaped bone, in which the hyomandibular facet is into the upper third of the anterior edge, the ventral edge covers part of the subopercle bone, and the lateral surface is smooth except for numerous dome-like folds present in the middle. The subopercle is a flat, long, and crescent shape-like structure, with a prominent anterior ascending process, a convex ventral edge, and probably a straight dorsal edge. Possibly, two disarticulated and scattered branchiostegal rays are preserved (Figure 3).
Pectoral girdle and fin. In MUZ-3912 preserves only fragments of cleithrum and possible posttemporal bones. A stout fragment of the ascending limb of the cleithrum is recognizable below the opercle. The posttemporal bone seems to be represented by a large and flat bone that covers part of the occiput of this fish (Figure 3).
Although incomplete and scattered, numerous elements of the left and right pelvic fins are present. At least seven proximal pectoral radials from both fins are preserved between the cleithrum and the pectoral rays; the largest is rectangular and massive in appearance, and the remaining tend to be ovoid and thinner. The proximal remains of at least 14 rays, also of both fins, are present; among these, the most anterior ones are in life position. The pectoral ray one is especially robust and thick; at least, it is twice as wide as ray two. These rays are unsegmented nor branched near the base (Figure 3).
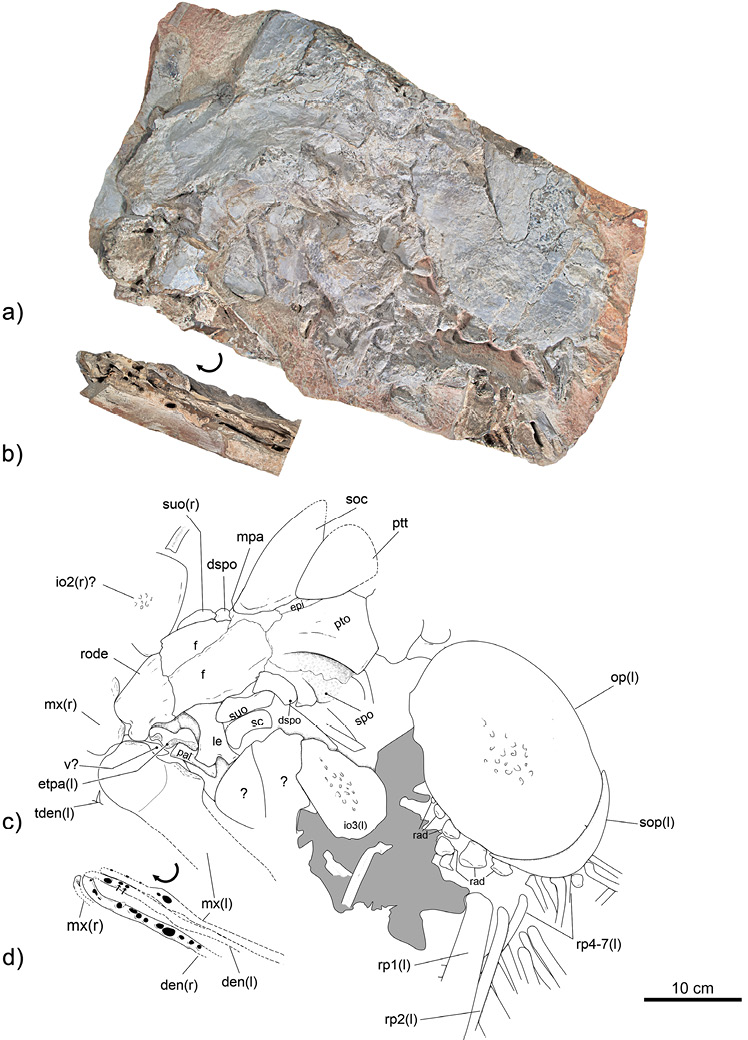
Figure 3. MUZ-3912, a Xiphactinus audax specimen from the Piedritas site, northern Coahuila, Mexico. a) Photography under white light; b) Section of the jaws exposed in the lateral surface of the slab, showing osseous remains of the maxilla and dentary bones as well as molds and teeth in cross section; c) idealized line drawing based on a); d) idealized line drawing based on b). Abbreviations: den, dentary; dspo, demosphenotic; epi, epioccipital; etpa, ethmopalatine; fr, frontal; io, infraorbital; le, lateroethmoid;mpa, mesoparietal; mx, maxilla; op, opercle; pal, palatine; pto, pterotic; ptt, posttemporal; rode, rostrodermethmoid; rad, radials; rp, ray of the pectoral fin;sc, sclerotic; soc, supraoccipital; sop, subopercle; spo, sphenotic; suo, supraorbital; t, tooth; v, vomer; rounded black patches on dentaries and maxillae show alveoli; black arrows on the left dentary show crown teeth in cross-section.
DISCUSSION
Although MUZ-3912 is an incomplete specimen and represents only the anterior part of a fish; it has sufficient morphological features to support its inclusion within the order Ichthyodectiformes, the suborder Ichthyodectoidei, the family Ichthyodectidae, and the genus Xiphactinus. This specimen is well established as a member of the order Ichthyodectiformes because it shows two of those diagnostic characteristics already identified by Bardack and Sprinkle (1969) and Patterson and Rosen (1977:115), these include the presence of (Figure 3): 1) the ethmopalatine bones forming the floor of the nasal capsule; and 2) a single row of teeth in the jaws. Today the first of these characters represents an unquestionable synapomorphy of the order Ichthyodectiformes. Other diagnostic characteristics of the order not observed in MUZ-3912 are the first three or four uroneurals extending anteroventrally covering the entire lateral surface of the first, second, or third preural centra; coracoids enlarged ventrally, meeting each other in a midventral coracoid symphysis; and the anal and dorsal fin opposite and remote in the posterior part of the body.
The history of the suborder Ichthyodectoidei is somewhat complex; it was vaguely erected by Romer (1966) as part of the Osteoglossomorpha to include all the allies of the genus Ichthyodectes, previously named as family Ichthyodectidae by Crook (1892), plus the members of the family Saurodontidae, named by Cope (1870) to Saurodon and Saurocephalus Harlan, 1824. Bardack and Sprinkle (1969) erected the Order Ichthyodectiformes including the Romer’s Ichthyodectoidei, considered all the non-saurodontid Jurassic and Cretaceous Ichthyodectoidei taxa as part of the family Ichthyodectidae. Regarding the Bardack and Sprinkle’s ichthyodectoidei taxa, Patterson and Rosen (1977: 115) erected the suborder Allothrossopoidei to include Allothrissops Nybelin, 1964; later, Occithrissops Arratia, Scasso, and Kiessling, 2004, was allocated more recently into this clade.
Although currently, the phylogenetic hypotheses of the Ichthyodectiformes are somewhat different, in particular in the naturalness and interrelationships of the suborder Ichthyodectoidei (see Taverne, 1986; Maisey, 1991; Stewart, 1999; Alvarado-Ortega, 2005; Taverne and Chanet, 2000; Cavin et al., 2013; Cavin and Berrell, 2019, Baños-Rodríguez et al., 2020); it is possible to point out that these fishes seem to be well naturally delimited into two groups, the first represented by the Allothrossopoidei known only by Jurassic species and the other (widely recognized as ichthyodectoidei) that includes Thrissops as the only Jurassic representative plus all the more derived species that developed during the Cretaceous. Different authors have suggest that the members of the suborder Ichthyodectoidei differ from the Allothrissopoidei by presenting more derived features on the head (Maisey, 1991; Alvarado-Ortega, 2005; Cavin et al., 2013; among others); continuing with the numbering of the previous paragraph, these features include: 3) supraoccipital and epioccipital bones elongated anteriorly to the level of the posterior edge of the orbit; 4) presence of a well-developed, high, and triangular supraoccipital crest; 5) the articular head of the palatine well-developed, massive, and malleolus-shaped. The inclusion of MUZ-3912 within Ichthyodectoidei is supported because it also presents all these features (Figure 3). It is noticeable that Dugaldia emmilta Cavin and Berrell, 2019, from the Albian deposits of Australia, lacks characters 3 and 4. Other features of this suborder not observed in MUZ-3912 probably due to its bad preservation, include the presence of posterior ridges in the epioccipital bones, and the membranous sutures of the ethmopalatine, and the presence of a basal sclerotic with serrated edges.
Among Cretaceous ichthyodectoidei fishes, Xiphactinus shoes a distinctive mixture of features of the supraoccipital crest, the dorsal view of the rostrodermethmoid bone, the shape of the maxilla, the distribution and size of the mandibular teeth, as well as in the proportions of the first pectoral rays. Continuing with the numbering of the previous paragraphs, these features include: 6) the supraoccipital crest involves a small participation of the mesoparietal bone, which posteriorly rises and forms the anterior part of this crest [Although this feature is restricted to those species belonging to Xiphactinus, Ichthyodectes and Gillicus (Bardack, 1965) as well as those of Saurodon, Saurocephalus, and Prosaurodon Stewart, 1999; in other Cretaceous ichthodectiforms this crest is formed only by the supraoccipital bone, as in Cladocylus Agassiz, 1841; Chiromystus Cope, 1885; and Unamichthys]; 7) in dorsal view, the rostrodermethmoid shows a well-developed lateral processes projected anterosposteriorly, beyond the bone becomes wider and its posterior edge a is straight [Among ichthodectiforms, the dorsal view of this bone shows different shapes (Alvarado-Ortega, 2005, fig. 6.9; Blanco-Piñón and Alvarado-Ortega, 2007, fig. 9); its posterior edge may have a deep medial notch or be entirely straight as in Cladocyclus; its lateral processes may be well developed and clearly extended anterolaterally or poorly developed and hardly expanded laterally as in Gillicus; and the back half of the bone may be tapered as in Cladocyclus, have concave edges as in Ichthyodectes, or become more expanded behind the lateral processes as in Prosaurodon]; 8) teeth are irregularly sized and spaced in the jaw bones [Among the ichthyodectiforms the devolvement of teeth of irregular size is notorious in the lower jaw of numerous Cretaceous species; however, this condition is extremely expressed and expanded to maxillar and premaxillar teeth only in Xiphactinus. In other Cretaceous ichthyodectiforms the dentary teeth may be absent together with those of the maxilla and premaxilla as in Heckelichthys; the teeth are well-developed and rather regular in size and more evenly distributed as in Vallecillicthys multivertebratum and Ichthyodectes; or these are notoriously smaller as in Gillicus and Saurodon,]; and 9) the first ray of the pectoral fin is notoriously breadth, at least two times wider than the second ray [Cavin et al. (2013, char. 10, state 0) revealed this feature as a synapomorphy shared by Ghrisichthys Cavin. Forey, and Giersch, 2013, and Xiphactinus. Although the pectoral fin ray 1 is breadth in other ichthyodectiforms, its width is not so extreme]. Here, MUZ-3912 is identified as a Xiphactinus specimen because it shows these four distinctive generic characters (6-9).
In his review of Xiphactnus, Bardack (1965) recognized three nominal species in addition to the type species, X. audax from North America, these are X. australis (Woodward, 1894) from marine deposits of the Early Cretacoeus in Queensland, Australia; X. mantelli Newton, 1877, from Cenomanian-Santonian deposits of England, Poland, France, Belgium, and Czech Republic; and X. galtinus Newton, 1877, from the Albian-Cenomanian deposits at Kent, England. More recently, Lees and Bartholomai, 1987, reviewed the specimens of X. australis, removed them from Xiphactinus, and renamed them as the type series of their Cooyoo australis. Remarkably, Bardack (1965) was unable to identify morphological features to distinguish the European species indicated from X. audax; he maintained these as distinct species only because of their distant geographical distribution. So far, the validity of such European species is questionable.
The valid nominal species of Xiphactinus considered here mainly dental differences. In X. audax, the teeth are uniformly conical and so, their transversal sections are rounded and have no sharp edges or carinae. On the contrary, in X. vetus, the teeth are lateromedially compressed and exhibit anterior and posterior sharp edges, thus in the transversal section, these are remarkably oval. Here, MUZ-3912 is referred to as X. audax because its teeth are conical and rounded in the transversal section.
Vavrek et al. (2014) provided a comprehensive review of the age and geographic distribution of Xiphactinus throughout USA and Canada. Here, such a review is extended to all of America including the present finding of X. audax in Coniacian-Campanian deposits of Coahuila plus two previous reports of indeterminate specimens of this genus, one recovered from the Cenomanian limestones of the La Luna Formation, Trujillo, Venezuela (Carrillo-Briseño et al., 2012) and that from the Latest Maastrichtian deposits of the Salamanca Formation, in Chubut Province, Argentina (De Pasqua et al., 2020) (Figure 4).

Figure 4. Distribution of Xiphactinus in America during the Late Cretaceous, gray show emerged continental territories (modified from Vavrek et al., 2014, fig. 4).
CONCLUSIONS
The discovery of MUZ-3912 in the Coniacian-Campanian deposits of the Austin Group, at the site of Piedritas, Ocampo, Mexico, and its specific determination as Xiphactinus audax are significant for distinct reasons. On the one hand, this fossil confirms the presence of this species in Mexico, which had already announced by Maldonado-Koerdell (1956) but later denied by Alvarado-Ortega (2005). This fish and those seven ichthyodectiform genera previously found in different Mexican localities equal the generic diversity of this order in the rest of North America, making Mexico an important territory to understand the diversity and evolution of this clade. This specimen expands the geographical distribution of the species to the south, a little beyond the US-Mexico border, filling a geographical gap between North and South America where it was already expected to find this genus revealing the wide longitudinal distribution of Xiphactinus throughout America, up to near both poles.
ACKNOWLEDGEMENTS
This research was financially supported by the Universidad Nacional Autónoma de México through the DGAPA-PAPIIT projects IN 209017 and IN 110920. We are in debt to Mr. Juan Quintana Franco for his help in the recovery of fossils in the Piedritas and other near paleontological sites. Mr. Juan M. Contreras helped us with the photos included in this work. The manuscript was improved with the valuable suggestions provided by Dr. Segio Bogan and an anonymous reviewer.
REFERENCES
Agassiz, L., 1833–1844, Recherches sur les poissons fossiles: Neuchâtel et Soleure, 5 volumes.
Alvarado-Ortega, J., 1998, Ichthyodectiformes mexicanos –un análisis de su diversidad–, in Carranza-Castañeda O., Córdoba-Méndez, D.A. (eds.), Avances en Investigación, Paleontología de Vertebrados: Universidad Autónoma del Estado de Hidalgo, Instituto de Investigaciones en Ciencias de la Tierra, Publicación especial 1, 109-119.
Alvarado-Ortega J, 2004, Description and relationships of a new ichthyodectiform fish from the Tlayúa Formation, Puebla, Mexico: Journal of Vertebrate Paleontology, 24, 4, 802-813.
Alvarado-Ortega, J., 2005, Sistemática de los peces Ichthyodectiformes de la Cantera Tlayúa, Puebla, México: Mexico City, Mexico, Instituto de Geología, Universidad Nacional Autónoma de México, Doctoral Thesis, 302 pp.
Alvarado-Ortega, J., Brito, P.M., 2010, A new ichthyodectiform (Actinopterygii, Teleostei) from the Lower Cretaceous Marizal Formation, north‐east Brazil: Palaeontology, 53, 2, 297-306.
Alvarado Ortega, J., Porras Múzquiz H.G., 2015, Xiphactinus en México, in Reynoso V.H., Flores Mejía, P., Aguilar, F., Moreno Bedmar, J.A. (comp.), Programa y Resúmenes del XIV Congreso Mexicano de Paleontología: Melchor Múzquiz, Coahuila, México, Sociedad Mexicana de Paleontología, Paleontología Mexicana, 19-24 de octubre, 2015, volumen especial 1, 25.
Alvarado-Ortega, J., Porras-Múzquiz, H., 2009, On the occurrence of Gillicus arcuatus (Cope, 1875) (Pisces, Ichthyodectiformes) in Mexico: Boletín de la Sociedad Geológica Mexicana, 61, 215-224
Alvarado-Ortega, J., Blanco-Piñón, A., Porras-Múzquiz, H., 2006a, Primer registro de Saurodon (Teleostei: Ichthyodectiformes) en México: Revista Mexicana de Ciencias Geológicas, 23(1), 107-112.
Alvarado-Ortega, J., Garibay-Romero, L.M., Blanco-Piñón, A., González-Barba, G., Vega-Vera, F.J., Centeno-García, E., 2006b, Los peces fósiles de la Formación Mexcala (Cretácico Superior) en el estado de Guerrero, México: Revista Brasileira de Paleontologia, 9(3), 261-272.
Alvarado-Ortega, J., González-Rodríguez, K.A., Blanco-Piñón, A., Espinosa-Arrubarrena, L., Ovalles-Damián, E., 2006c, Mesozoic Osteichthyans of Mexico, in Vega, F.J., Nyborg ,T.G., Perrilliat, M.C., Montellano Ballesteros, M., Cevallos Ferriz S.R.S., Quiroz Barroso, S.A., (eds.), Studies on Mexican Paleontology, Topics on Geobiology, 24: Dordrecht, The Netherlands,Springer, 160-207.
Alvarado-Ortega, J., Ovalles-Damián, E., Blanco-Piñón, A., 2009, The fossil fishes from the Sierra Madre Formation, Ocozocoautla, Chiapas, Southern Mexico: Palaeontologia Electronica, 12, 2(4a), 1-22.
Alvarado-Ortega, J., Cantalice, K.M., Martínez-Melo, A., García-Barrera, P., Than-Marchese, B.A., Barrientos-Lara, J.I., Díaz-Cruz, J.A., 2020, Tzimol, a Campanian marine paleontological site of the Angostura Formation near Comit_an, Chiapas, southeastern Mexico: Cretaceous Research, 107, 104279.
Arratia, G., Scasso, R.A., Kiessling, W., 2004, Late Jurassic fishes from Longing Gap, Antarctic Peninsula: Journal of Vertebrate Paleontology, 24, 1, 41-55.
Bahrami, A., Yazdi, M., Moghaddam, H. V., Ghobadi, A., Murray, A. M., Brinkman, D., Parvaneh-Nejad Shirazi, M., Ataabadi, M. M., Kundrát, M., 2021, Early Cretaceous vertebrate and invertebrate fossils from Dariyan Formation, southern Iran: Historical Biology, 33(3), 387-402.
Baños Rodríguez, R.E., 2018, Taxonomía y Sistemática de los peces Ichthyodectiformes (Teleostei) de la Cantera Muhi (Albiano-Cenomaniano) de Zimapán, Hidalgo, México: Pachuca, Hidalgo, Mexico, Universidad Autónoma del Estado de Hidalgo, Master Thesis, 89 pp.
Baños-Rodríguez, R.E., González-Rodríguez, K.A., Wilson, M.V., González-Martínez, J.A., 2020, A new species of Heckelichthys from the Muhi Quarry (Albian–Cenomanian) of central Mexico: Cretaceous Research, 110, 104415.
Bardack, D., 1965, Anatomy and evolution of chirocentrid fishes: The University of Kansas Paleontological Contributions, Vertebrata, 10, 1-88.
Bardack, D., Sprinkle, G., 1969, Morphology and relationships of saurocephalid fishes: Fieldiana Geology, 16, 297-340.
Berrell, R.W., Alvarado-Ortega, J., Yabumoto, Y., Salisbury, S.W., 2014, The first record of the ichthyodectiform fish Cladocyclus from eastern Gondwana: a new species from the Lower Cretaceous of Queensland, Australia: Acta Palaeontologica Polonica, 59, 4, 903-920.
Blanco-Piñón A, Alvarado-Ortega J, 2005, Fishes from la Mula Quarries, a new Late Cretaceous Locality from the vecinity of Múzquiz, Coahuila, NE México, in Poyato-Ariza, F.J. (ed.), Fourth Internacional Meeting on Mesozoic Fishes –Systematics, Homology, and Nomenclature- Extended Abstracts: Miraflores de la Sierra, Madrid, España, UAM ediciones, 37-41.
Blanco-Piñón, A., Alvarado-Ortega, J., 2007, Review of Vallecillichthys multivertebratum (Teleostei: Ichthyodectiformes), a Late Cretaceous (early Turonian)" Bulldog fish" from northeastern Mexico: Revista Mexicana de Ciencias Geológicas, 24(3), 450-466.
Blanco, A., Cavin, L., 2003, New Teleostei from the Agua Nueva Formation (Turonian), Vallecillo (NE Mexico): Comptes Rendus Palevol, 2(5), 299-306.
Carrillo-Briseño, J., Alvarado-Ortega, J., Torres, C., 2012, Primer registro de Xiphactinus Leidy, 1870 (Teleostei, Ichthyodectiformes) en el Cretácico Superior de América del Sur (Formación La Luna, Venezuela): Revista Brasileira de Paleontologia, 15(3), 327-335.
Cavin, L., Berrell, R.W., 2019, Revision of Dugaldia emmilta (Teleostei, Ichthyodectiformes) from the Toolebuc Formation, Albian of Australia, with comments on the jaw mechanics: Journal of Vertebrate Paleontology, 39, 1, e1576049.
Cavin, L., Forey, P.L., Giersch, S., 2013, Osteology of Eubiodectes libanicus (Pictet & Humbert,) and some other ichthyodectiformes (Teleostei): phylogenetic implications: Journal of Systematic Palaeontology, 11(2), 115-177.
Cope, E.D., 1870, On the Saurodontidae: Proceedings of the American Philosophical Society, 11, 529-538.
Cope, E.D., 1871, On two extinct forms of Physostomi of the Neotropical region: Proceedings of the American Philosophical Society, 12, 52-55.
Cope, E.D., 1875, The vertebrata of the Cretaceous formations of the west: Report, US Geological Survey Territories, 2, 1-303.
Cope, E.D., 1885, A contribution to the vertebrate paleontology of Brazil: American Philosophical Society, Proceedings, 17, 176-181.
Crook, A.R., 1892, Über eingie fossile Knochenfische aus der mittlern Kreide von Kansas: Paleontographica, 39, 107-124 + 3 pls.
De Pasqua, J.J., Agnolin, F.L., Bogan, S., 2020, First record of the ichthyodectiform fish Xiphactinus (Teleostei) from Patagonia, Argentina. Alcheringa, 44(2), 327-331.
Dixon, F., 1850, The geology and fossils of the Tertiary and Cretaceous formations of Sussex: London, Longman, Brown, Green and Longmans, 422 pp. + 45 pl.
Felix, J., 1891, Versteinerungen aus der mexicanischen Jura und Kreide Formation: Palaeontographica, 37, 140-194.
Giersch, S., 2014, Die Knochenfische der Oberkreidezeit in Nordostmexiko: Beschreibung, Systematik, Vergesellschaftung, Paläobiogeographie und Paläoökologie: Germany, Ruprecht-Karls-Universität Heidelberg, PhD Thesis, 275 pp.
Giersch, S., Frey, E., Stinnesbeck, W., Gonzalez-Gonzalez, A.H., 2008, Fossil fish assemblages of northeastern Mexico: new evidence of mid Cretaceous Actinopterygian radiation, in Krempaska, Z. (ed.), 6th Meeting of the European Association of Vertebrate Paleontology: Slovak Republic, Museum of Spiš, Spišska Nova Ves, Volume of Abstracts, 43-45.
Harlan, R., 1824, On a new fossil of genus of the order Enalio Sauri (of Conybeare). Philadelphia: Journal of Academy of Natural Science, 3, 331-337 + 1 pl.
Hays, I., 1830, Description of a fragment of the head of a new fossil animal, discovered in a marl pit, near Moorestown, New Jersey: Transactions of the American Philosophical Society, Series 2, 3(18), 471-477+ pl. 16.
Heinz, R., 1932, Aus der neue Systematik der Inoceramen: Mitteilungen aus dem Mineralogisch-Geologischen Staatsinstituts in Hamburg, 13, 1-26.
Jaekel, O., 1909, Beiträge zur Geologie von Kamerun. X. Fischreste aus der Mamfe Shiefern: Köriglische Preussische Geologische Landesantstalt, Abhandlung, new series, 6, 392-398.
Jordan, D.S., 1910, Description of a collection of fossil fishes from the bituminous shales of Riacho Doce, State of Alagoas, Brazil: Anales of Cargenie Museum, 7, 1, 22-34.
Kim, H.M., Chang, M.M., Wu, F., Kim, Y.H., 2014, A new ichthyodectiform (Pisces, Teleostei) from the Lower Cretaceous of South Korea and its paleobiogeographic implication: Cretaceous Research, 47, 117-130.
Lees, T., Bartholomay, A., 1987, Study of a Lower Cretaceous actinopterygian (Class Pisces) Cooyoo australis from Queensland, Australia. Memoirs of the Queensland Museum, 25(1), 177-192.
Leidy, J., 1870, Remarks on ichthyodorulites and on certain fossil Mammalia: Proceedings of Academy on Natural Science of Philadelphia, 22, 12-13.
Maisey, J.G., 1991, Cladocyclus Agassiz, 1841, in Maisey, J.G. (ed.), Santana Fossils. An Illustrated Atlas: Neptune City, T.H.F. Publications, Inc., 190-207.
Maldonado-Koerdell, M., 1956, Peces fósiles de México III. Nota preliminar sobre peces del Turoniano superior de Xilitla, San Luis Potosí, México: México, Ciencia, 16, 1-3, 31-36.
Marsh, O.C., 1872, Notice of a new and remarkable fossil bird: The American Journal of Science and Arts, series 3 and 4, 19-24, 344 pp.
Müller, J., 1845, Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische: Archiv für Naturgeschichte, 11, 1, 91-141.
Newton, E.T, 1877, On the remains of Hypsodon, Portheus and Ichthyodectes from British Cretaceous strata, with descriptions of a new species: The Quarterly journal of the Geological Society of London, 33, 505-529 + 1pl.
Nybelin, O., 1964, Versuch einer taxonomischen revision der jurassischen Fischgattung Thrissops Agassiz: Göteborgs K. Vetensk.-o. Vitterh.-Samh. Handl. 6, ser. B, 9, 4, 1-44.
Patterson, C., Rosen, D.E., 1977, Review of ichthyodectiform and other Mesozoic teleost fishes, and the theory and practice of classifying fossils: Bulletin of the American Museum of Natural History, 158, 2, 1-172.
Porras-Múzquiz, H.G., Chatterjee, S., Lehman, T.M., 2014, The carinate bird Ichthyornis from the Upper Cretaceous of Mexico: Cretaceous Research, 51, 148-152.
Riquelme, F., Alvarado-Ortega, J., Ruvalcaba-Sil, J.L., Aguilar-Franco, M., Porras-Múzquiz, H., 2013, Chemical fingerprints and microbial biomineralization of fish muscle tissues from the Late Cretaceous Múzquiz Lagerstätte, Mexico: Revista Mexicana de Ciencias Geológicas, 30(2), 417-435.
Romer, A.S., 1966, Vertebrate Paleontology: Chicago, IL, USA, University of Chicago Press, X+478 pp.
Schaeffer, B., Patterson, C., 1984, Jurassic fishes from the western United States, with comments on Jurassic fish distributions: American Museum Novitates, 2796, 1-86.
Sowerby, J., 1814, The Mineral Conchology of Great Britain, or coloured figures and descriptions of those remains of testaceous animals or shells which have been preserved at various times and depths in the earth: London, Benjamin Meredith, 242 pp+vii.
Springer, V.G., 1957, A new genus and species of elopid fish (Laminospondylus transversus) from the Upper Cretaceous of Texas: Copeia, 1957(2), 135-140.
Stewart, J.D., 1999, A new genus of Saurodontidae (Teleostei: Ichthyodectifonnes) from the Upper Cretaceous rocks of the Western Interior of North America, in Arratia, G., Schultze, H.-P. (eds.), Mesozoic Fishes 2: München, Systematics and Fossil Record, Verlag Dr. F. Pfeil, 335-360.
Stinnesbeck, W., Ifrim, C., Schmidt, H., Rindfleisch, A., Buchy, M.C., Frey, E., González-González, A.H., Vega, F.J., Cavin, L., Keller, G., Smith, K.T., 2005, A new lithographic limestone deposit in the Upper Cretaceous Austin Group at El Rosario, county of Múzquiz, Coahuila, northeastern Mexico: Revista Mexicana de Ciencias Geológicas, 22, 3, 401-418.
Taverne, L., 1986, Ostéologie et affinités systématiques de Chirocentrites vexillifer du Crétacé supérieur de la Mésogée Eurafricaine. Considérations sur la phylogénie des Ichthyodectiformes, poissons téléostéens du Jurassique et du Crétacé: Annales de la Société Royale Zoologique de Belgique, 116(1), 33-54.
Taverne, L., 2008, of the new genus Heckelichthys (Teleostei, Ichthyodectiformes)-A new visit inside the ichthyodectid phylogeny: Bulletin de l'institut Royal des Sciences Naturelles de Belgique Sciences de la Terre, 78, 209-228.
Taverne, L., Chanet, B., 2000, Faugichthys loryi n. gen., n. sp. (Teleostei, Ichthyodectiformes) de l'Albien terminal (Crétacé inférieur marin) du vallon de la Fauge (Isère, France) et considérations sur la phylogénie des Ichthyodectidae: Geodiversitas, 22(1), 23-34.
Than-Marchese, B.A., Alvarado Ortega, J., 2011, Los peces fósiles cenomanianos de Chiapas, México, y su importancia taxonómica y biogeográfica, in IV Congreso Latinoamericano de Paleontología de Vertebrados: San Juan Argentina, Asociación Paleontológica Argentina, CD de resúmenes, resumen 103.
Vavrek, M.J., Murray, A.M., Bell, P.R., 2014, Xiphactinus audax Leidy 1870 from the Puskwaskau Formation (Santonian to Campanian) of northwestern Alberta, Canada and the distribution of Xiphactinus in North America: Vertebrate Anatomy Morphology Palaeontology, 1, 89-100.
Villaseñor, A.B., Silva-Martínez, L.E., Olóriz, F., Blanco, A., Alvarado Ortega, J., 2006, Nuevo registro de peces Jurásicos en México, in Memorias del X Congreso Nacional de Paleontología: Mexico City, Mexico, Sociedad Mexicana de Paleontología, p. 137.
Woodward, A.S., 1894, On some fish-remains of the genera Portheus and Cladocyclus from the Rolling Downs Formation (lower Cretaceous) of Queensland: The Annals and Magazine of Natural History, series 6, 14, 444-447 + 1 pl.
Yabumoto, Y., Hirose, K., Brito, P.M., 2018, A new ichthyodectiform fish, Amakusaichthys goshouraensis gen. et sp. nov. from the Upper Cretaceous (Santonian) Himenoura Group in Goshoura, Amakusa, Kumamoto, Japan: Historical Biology, 32, 3, 362-375.