ABSTRACT
The Calera cave system in San Luis Potosí, Mexico, with a large Pleistocene megafaunal assemblage is described. Hundreds of fossil bones have been found throughout the 1324 m of mapped galleries. The fossils are well preserved resulting from partial permineralization by manganese oxide. A preliminary taxonomic identification of the recovered specimens indicates Equus sp. is the most abundant taxon. Other groups include mammoths, gomphotheres, cervids, bovids, camelids, sloths, glyptodonts, felids, canids, lagomorphs, and testudines. It has been estimated that the cave would have started to form between 1.17 and 0.29 Ma. Since the fossils are found on top of or form part of the fluvial sedimentary deposits inside the cave passages, fossil deposition is much younger and probably occurred during the latest stage of cave formation and the fauna represents a time-averaged accumulation. A few specimens are covered with secondary calcite incrustations. The locality described in this study stands out because it contains one of the largest Pleistocene megafaunal assemblages of the region.
Key words: fossil; megafauna; cave; equid; mammoth; gomphothere; sloth, Pleistocene; Calera cave system; Sierra de El Abra; San Luis Potosí; Mexico.
RESUMEN
Se describe el sistema Calera en San Luis Potosí, México, que contiene un gran conjunto de fósiles representantes de la megafauna pleistocénica. Cientos de huesos fósiles se pueden encontrar a lo largo de los 1324 m de galerías mapeadas. Por lo general están en relativo buen estado de conservación gracias a haber sido permineralizados por oxido de manganeso. Una revisión taxonómica preliminar muestra que los restos más abundantes son miembros del género Equus. Otros grupos presentes son mamuts, gonfoterios, cérvidos, bóvidos, camélidos, perezosos, gliptodontes. félidos, canidos, lagomorfos y tortugas. Se estima que la cueva se empezó a formar entre 1.17 y 0.29 Ma. Debido a que los fósiles se encuentran en la superficie o formando parte de las capas superiores de sedimentos fluviales (gravas y arenas) depositados dentro de la cueva, el depósito de los fósiles es mucho más joven y probablemente ocurrió en los últimos episodios en la formación de la cueva; Algunos ejemplares han sido cubiertos por incrustaciones calcáreas. La relevancia de esta localidad está dada por ser una de las mayores concentraciones de restos de megafauna pleistocénica en la región.
Palabras clave: fósil; megafauna; cueva; mamut; gonfoterios; perezoso; Pleistoceno; sistema Calera; Sierra de El Abra; San Luis Potosí; México.
Manuscript received: May 2, 2024
Corrected manuscript received: August 25, 2024
Manuscript accepted: August 27, 2024
Published Online: December 1, 2024
INTRODUCTION
The Sierra de El Abra, in northeast Mexico, spans the states of Tamaulipas and San Luis Potosí and is the front range of the Sierra Madre Oriental of México (Supplementary Figure S1). Mostly composed of Cretaceous limestone, it is dominated by a karstic landscape formed by the groundwater dissolution of limestone with over 200 caves explored in the El Abra region. The geology of the Sierra de El Abra has been studied intensively due to the oil deposits that occur in similar rocks buried under the coastal plain (Boyd, 1963; Carrillo-Bravo, 1971; Enos, 1974; Aguayo-Camargo, 1998). Caves in this area have a long history of exploration. Much of the fieldwork in this region was initially driven by an interest in Astyanax, the Mexican blind cavefish (Bonet, 1953; Russell & Raines, 1967; Mitchell et al., 1977; Ford, 2000; Fish, 2004; Elliott, 2015; Espinasa and Espinasa, 2015; Elliott, 2018; Keene et al., 2015). One of us (LE) has led new cave explorations, resulting in the discovery and description of new caves (Espinasa et al., 2020). During the exploration of one of them, the Calera cave system, a large assemblage of fossilized bones was noticed. The cave is prone to periodic flooding and most fossils have been transported down passages during flood events. Furthermore, it is visited occasionally by locals, who might trample over the remains or may remove some specimens. In consideration of the above, four expeditions were carried out to complete the cave survey, study these fossils, and register their location.
Polaco & Butrón (1997) initially reported 14 caves in México where fossil vertebrate remains had been found. Six years later, Arroyo-Cabrales & Polaco (2003) recounted 18 different cave sites and described their Pleistocene fauna, which represents more than half of the known taxa of Pleistocene fossil mammals of Mexico. More recently Ferrusquía-Villafranca et al. (2017) and Bonilla Díaz (2018) mentioned 27 different cave sites with Pleistocene (Rancholabrean) materials. The ongoing collaborations between cavers and paleontologists are bound to further increase this list (McDonald, 2020).
Despite the intense work the caves of the Sierra de El Abra have inspired, Pleistocene fossil remains have previously only been reported for one cave, Cueva de El Abra (Dalquest & Roth, 1970; Holman, 1970), but no megafauna was identified as these authors concentrated on smaller fossils, mostly of rodents, bats, amphibians, and reptiles.
Of the many paleontological sites known in San Luis Potosí, only ten have Pleistocene fauna: El Cedral (Lorenzo and Mirambell, 1981; Polaco-Ramos, 1981; Álvarez & Polaco, 1982; Alberdi et al., 2003, 2014), Laguna Media Luna (Hernández Junquera, 1977), Brechas Coloradas (Freudenberg, 1919, 1921; Pichardo Del Barrio, 1960), Rancho Peotillos (Silva-Barcenas, 1969; Ferrusquía-Villafranca et al., 2016), Hacienda La Tinaja (Arellano, 1951), Laguna de Las Cruces (Álvarez, 1977; Perez-Crespo et al, 2012, 2013), La Presita (Polaco & Butrón, 1997; Bonilla Díaz, 2018), Mina San Antonio (Torres-Martínez, 1995; De Anda, 2009), La Presita Blanca (Marín-Leyva et al., 2018) and Vichinchijol, Ebano (Arroyo-Cabrales et al., 2007; Tapia-Ramírez et al., 2013; Polaco y Aguilar, 1999).
Nearby open air and cave sites with Pleistocene fossils in Tamaulipas are also scarce: Ejido San Lázaro, Municipio de Villagrán, 80 km N of Ciudad Victoria (Reynoso & Montellano-Ballesteros, 2004), the previously mentioned Cueva de El Abra (Dalquest & Roth, 1970; Holman, 1970), and El Salitrillo, near Villa Xicoténcatl (Riva-Palacio, 1969).
The locality described in this study (Supplementary Figure S1) stands out because, given the unique conditions of this cave, it resulted in the concentration of large vertebrate fossil remains. Thus, we describe here one of the largest Pleistocene megafaunal assemblages of the region.
GEOLOGY AND GEOMORPHOLOGY
The Sierra de El Abra is primarily composed of limestones of the El Abra (or El Doctor) Formation, deposited as part of a rudist reef complex which ringed the Valles-San Luis Potosí calcareous platform during the mid-Cretaceous (Albian to Turonian) (Carrillo-Bravo, 1971). The steep eastern front of the range represents the reef escarpment and is composed of muddy rudist banks and the much more abundant inter-reef bioclastic debris and mud (Bonet, 1952; Bonet, 1963). The rest of the El Abra limestone is composed of the backreef or platform facies, with a variety of lithofacies including calcilutites and calcarenites with miliolids and toucasia rudist biostromes. This facies is where the cave under study, the Calera system, developed (Supplementary Figure S2). During the Late Cretaceous, the deposition of calcareous sediments slowly changed to terrigenous sediments (shales) of the Agua Nueva, San Felipe, and Mendez formations, which progressively covered the platform and reef. In the immediate area of Calera system, the Agua Nueva Formation was not deposited, and the Mendez Formation has been eroded away, in such a way that the northernmost entrances of Calera system are at the contact between the San Felipe shales (Supplementary Figure S2c, S2d, S2e and the limestone (Supplementary Figure S2a, S2b, S2e).
During the early Paleocene, the area was folded and uplifted. While the Sierra de El Abra and surrounding area was not very intensively deformed, a series of four small anticlinal ridges, somewhat parallel to the trend of the mountain range can be discerned in the study area (Supplementary Figure S3). As soon as they were uplifted, the shales began to erode, and the exposed limestone developed into a high-relief karst terrain, formed by the dissolving action of slightly acidic groundwater moving along both joints (vertical fractures) and horizontal bedding planes (Fish, 2004; Elliot 2018).
In the areas with shales, stream flow is on the surface of the land, as the shales are impervious to water. However, where limestone is exposed, water sinks underground through the epikarst and there are no surface streams. Stream capture into the underground occurs when the overlying San Felipe and Mendez shales are eroded away, and streams can invade the underlying El Abra limestone. The El Abra limestone probably was exposed first along high ridges before the present-day swallets formed in the lowlands near Ciudad Valles (Fish, 2004; Elliot, 2018).
KARST EVOLUTION AND GEOHYDROLOGY
The Calera system and the related Sótano del Toro developed as a capture of a small stream along one of the N-S synclinal valleys that form the western flank of the southern Sierra de El Abra (Supplementary Figure S3). Mitchell et al. (1977) and Fish (2004) describe numerous instances where surface streams on the western flank of the central Sierra de El Abra, flowing in a north-south direction over San Felipe shales, were captured underground as soon as the deepening of the surface drainage by stream erosion reached the underlying limestones. These authors also argue that the canyon depth upstream of the sink can be indicative of the relative age of the cave. The stream entering the Calera system has not yet formed any canyon, suggesting this cave is very young.
Most of the major caves of the western flank of the central Sierra de El Abra (Tigre, Roca, Arroyo, Sabinos, Tinaja, Montecillos, Pichijumo, Jos, Piedras, Palma Seca and Curva) originated as stream captures, and probably drain southeast, across the range, towards the large resurgence of Nacimiento del Río Choy (Fish, 2004; Elliot, 2018). The southern Sierra de El Abra (south of the Highway 70 pass) also has at least one stream capture cave, Cueva Chica, which drains south towards Cueva Chiquitita and the Rio Tampaón (Supplementary Figure S4). Although the final destination of the water that flows through Calera system is not known, it is suggested that it surfaces at a temporary spring on the bed of an arroyo southwest of the cave.
METHODS
Exploration and study of the Calera cave system is both technically challenging and strenuous. The three main entrances to the cave system are vertical pitches of 12, 6 and 5 m, all of which require knowledge of vertical caving single-rope techniques (Supplementary Figure S5). The average height of the cave passages is 1.2 m, with many long sections requiring crawling on or over muddy gravel and boulders (Figure 1a) and/or sharp rocks (Figure 1b), and sometimes through pools of water (Figure 1c) up to 50 cm deep and with only 20 cm of breathing air space. Two particularly long flat-out belly crawls are less than 30 cm between ceiling and floor (Figure 1a and 1d). These conditions required the use of gloves, kneepads, helmets, harnesses, and caving lamps. Search for fossils in the deep pools was done with snorkel, mask, and a scuba diving light. In dry passages, visual observation without digging was performed. It is likely that removal of the finer sediments through excavation could expose more fossil material, especially microfauna.

Figure 1. a-d) Typical low passages in Sistema Calera. a) Looking into Arrastre Lodoso, upstream lower level of Dos Soyates. b) Upper-level downstream crawlway of Dos Soyates. c) Upstream lower level of Dos Soyates. d) Arrastre Seco connection crawl between Sótano del Toro 2 and Dos Soyates. e-h) Coatings on the fossils. e) Both the fossil (red arrow) and many of the pebbles are coated in a black, metallic luster of manganese oxide. f) Long bone fragment with slight manganese coating. Notice that it did not penetrate the bone, as can be seen where it is broken. Also, the breaking occurred after the manganese-coating event. g) Sloth vertebra completely permineralized in manganese-oxide. h) Calcite concretions growing on top of the manganese-oxide coating.
Cave entrances to Calera system were located using the GPS integrated on a Samsung S21FE and by using the Geo Tracker app from geo-tracker.org. The cave was mapped using the methods described in Ellis (1976), Dasher (1994) and Day (2002), by setting permanent stations along the cave passage, between which the distance was measured to the nearest millimeter with a Disto A5 laser rangefinder. Azimuth was measured to the nearest ½° using a Suunto KB-14 Compass, and inclination was measured to the nearest ½° using a Suunto PM-5 Inclinometer. At each survey station, distances to the left and right wall, ceiling and floor were also measured with the Disto A5. Simultaneously with the recording of the previous data, a scaled and properly oriented sketch of the cave passage was made on a hand-held notebook, in which passage shape detail, sedimentary fills, flowstone and calcite decorations, and any other data of interest was plotted. Survey data was processed using the Compass and CaveXO software to generate plan and profile views of the cave. Particularly detailed sketches were made of the areas of the cave where fossils were found, specifying their relative locations.
Using a Samsung S21FE camera, significant fossils were imaged in situ, identified with a 1-inch-wide orange flagging tape with an inventory number/code. The location of significant fossils was registered on the map. Rescued fossils were collected in bags, with their number/code attached, and sealed to prevent sudden drying of the fossils. The specimens were collected under permit # 401.1S.3-2024/138 from INAH. Many specimens were left untouched because of their redundancy (e.g., equid-teeth or long bone fragments), requiring excavation to extract, or were considered too fragile or fragmentary to be identifiable to element or taxon.
The fossils were cleaned with tap water and a soft toothbrush to eliminate the mud covering them. They were slowly dried afterwards at room temperature. Once dried, all samples were photographed using a Nikon Z9 camera, with a 105-macro lens, lighted with two SB910 flashes synchronized with a SU 800 transmitter, and afterwards edited using Lightroom software. High resolution copies of these images are available to any interested researcher contacting the first author or the corresponding author.
Given that the main goal of this study was the description of the cave, no attempt for carbon dating was made given that the specimens’ fossilization under water suggests the leaching out of organic carbon from the samples. Future studies will corroborate the validity of this assumption.
Preliminary and tentative identification of the fossils was made through comparisons with published materials. No attempt was made to reach the species level as this will be the purpose of future work. Samples were grouped as morphospecies. Variability within these groups is limited to measurements made with a Vernier caliper of the length and width of tooth cusps. All fossil samples have been preserved and are kept in the custody of the Sociedad Mexicana de Exploraciones Subterráneas A.C. (SMES). Samples were given a SMES collection number (S1-S92) and an INAH registration number (3380PF 1-67).
RESULTS
Age of the cave system
Espinasa & Espinasa (2015) proposed a model for dating caves in the Sierra de El Abra based on when they were first exposed by erosion of the overlaying impervious layers. Given that the cave entrance is at an altitude of 92 masl and that of the base level is at 48 masl, they reached the conclusion that the Calera cave system would have started to form between 1.17 and 0.29 Ma.
According to Mitchell et al. (1977), the relative age of a stream sink in the west flank of the El Abra can be approximated depending on the depth of the canyon excavated in the El Abra and the height of the headwall at the entrance. The intermittent stream that enters Sótano del Toro during rainstorms has not excavated any perceptible canyon and the doline edge is right at the contact between the San Felipe shales and the underlying El Abra limestone. This would indicate this is an extremely young and recent stream capture, supporting the aforementioned age estimate. Nevertheless, the presence of an older, upper, and originally phreatic level (Figure 1a, 1b), which was later vadosely eroded before being abandoned in favor of the presently active lower level, which in turn is being pirated by a newer and even lower level (Figure 1c) as indicated by at least one and probably two episodes of water table lowering that have occurred since the cave’s inception. Vertebrate fossils are found throughout all passages that experience flooding, both phreatic and vadose, suggesting that the age of the primary accumulation of the fossils corresponds to the final and current erosive stage of the cave. Thus, the fossils are significantly younger than 1.17–0.29 Ma, which is supported by their apparent Rancholabrean age (ca. 240,000 to 11,000 years BP) based on the vertebrate remains.
Cave description
Sótano del Toro was originally explored in 1969 and described by Elliot (2018). Sótano del Toro 2 and Sótano Dos Soyates were partially explored in 2019 and described by Espinasa et al. (2020), who renamed Dos Soyates as cueva Calera. In December 2022 and January 2023, the cave was completely surveyed by the authors for a total length of 1,324 m, establishing the connection between Dos Soyates and Sótano del Toro 2 and discovering a new entrance, Sótano del Soyate Solitario (Supplementary Figure S5e).
Sótano del Toro (21°53’49”N, 98°55’52”W, 103 masl) is at the northern side of a doline 25 m in diameter and 6 m deep. During the rainy season a stream reaches this doline from the north and forms a cascade straight into the Sótano (Supplementary Figure S6). The access into the cave is a steep funnel that drops 8 m to a sump pool 2 m long by 0.5 m wide. It is thought that this sump pool connects through underwater galleries with the sump at the bottom of Sótano Toro 2. There is evidence from mud and debris that during flood events, the water backs up and the doline can be filled almost completely.
The Calera system proper has three known entrances (Supplementary Figure S7): Sótano del Toro 2, Sótano de los Dos Soyates, and Sótano del Soyate Solitario. Sótano del Toro 2 is a 12 m pitch, which is less than 0.5 m in diameter for the first 4 m and drops straight into a deep pool (Supplementary Figure S8a). This is probably the downstream end of the sump from Sótano del Toro (Supplementary Figure S7). A relatively roomy decantation chamber follows, where mud deposits several meters deep are abundant. Fossils were observed under the fine silts of these deposits where larger material such as rocks are present. The cave then separates into a lower, active level, and an upper, fossil passage. The lower level soon becomes a sump, but its continuation joins the lower level through the Dos Soyates entrance at the Llac Lluis. While this lower, active level is a typical phreatic passage and shows evidence of complete and severe flooding, the upper level, although originally also phreatic, shows that at some point subsequent vadose erosion carved a small canyon on the floor of the passage, before the water was diverted towards the lower level (Figure 1c). During extreme flood events the water level reaches the upper level. Most pools in the lower level have blind Astyanax cavefish, but none were seen in the few pools in the upper level.
Fossil-bearing sites
Fossils were found in every passage, but their concentration diminishes away from the entrance room of Sótano del Toro 2 (Supplementary Figure S7). They were found on the surface of or as part of fluvial (gravel and sand) sedimentary deposits inside the cave passages. In the lower, active level, the progressive diminishing in abundance is less intense, with fossil remains having been found all the way to the furthest final sump downstream to the south. This evidence ongoing transportation of fossil remains during flood events, moving remains that were originally present in Sótano del Toro and Sótano del Toro 2 farther into the cave. As would be expected from a site where remains are actively remobilized and transported by water throughout the cave, we did not find any clear sorting of fossils by taxonomy. Most associations consisted of similarly sized remains as documented by Voorhies (1969) in other water transported assemblages. Numbered red dots found within maps in this manuscript show the position of individual fossils. As mentioned in the methods section, high resolution images as well as their taxonomic assignment are available to any interested researcher contacting the first author or the corresponding author.
Northern portion
Both the Sótano del Toro and Sótano del Toro 2 entrance pits drop directly into deep pools (Supplementary Figures S6d and S8a), which are probably connected through a sump. During floods, this sump overflows and fills the large chamber of Sótano del Toro 2 to a depth of at least 3 m, before it can flow downstream towards the downstream sump (Supplementary Figure S8). This entrance chamber serves as a decantation pool, as indicated by the thick mud deposits. Fossils were found directly underwater in the main pool (Supplementary Figure S8c), on the surface of the talus slope near the water, and in the passage leading to the downstream sump (Supplementary Figure S8d). In this northern portion of the cave is where some of the largest and most complete remains were found. These included the maxilla of an equid and the mandible of a cervid, with most of their teeth still attached. There were also isolated teeth of two species of Proboscidea as well as part of a shoulder girdle. The femur of a Xenarthran was also located in this area. We presume that many other fossils could be found in the flooded connection with Sótano del Toro, under the thick mud deposits of the talus slope on the north side of the room, and in the Arrastre Seco connection crawlway with Dos Soyates.
Along the upper level, sediments and fossils are concentrated above the connection crawlway with the Toro 2 Chamber (Supplementary Figure S8d). The crawlway itself probably also has other fossils embedded in the sediments, but none were noticed along the path used to traverse it, as the passage height is just 30 cm (Figure 1d). During severe floods, water must push sediments up the crawlway and into the upper level, where the sediment load, including the fossils, is deposited.
Central portion
From Sótano del Toro 2 chamber, water flows downstream via a sump to Llac Lluis, where some large bones were found. The water enters a second sump which connects to the upstream lake of the lower level of Dos Soyates (Figure 2k). After this section the galleries go upward, creating another decantation pool. When severe flooding occurs, water fills up the Arrastre Lodoso and flows into the Dos Soyates lower level, but rarely has the energy to push fossils up El Pozo (Figure 2g). Water has pushed small boulders, sediments, and many fossils into a gravel embankment, creating one of the largest accumulations of fossils in the entire cave system (Figure 2b–2f). Close to 300 individual remains have been recorded in this embankment, but most of them were left in situ. Fossils that were transported by the water beyond El Arrastre Lodoso passage are found in progressively smaller quantities throughout the lower levels of the Dos Soyates section of the cave.
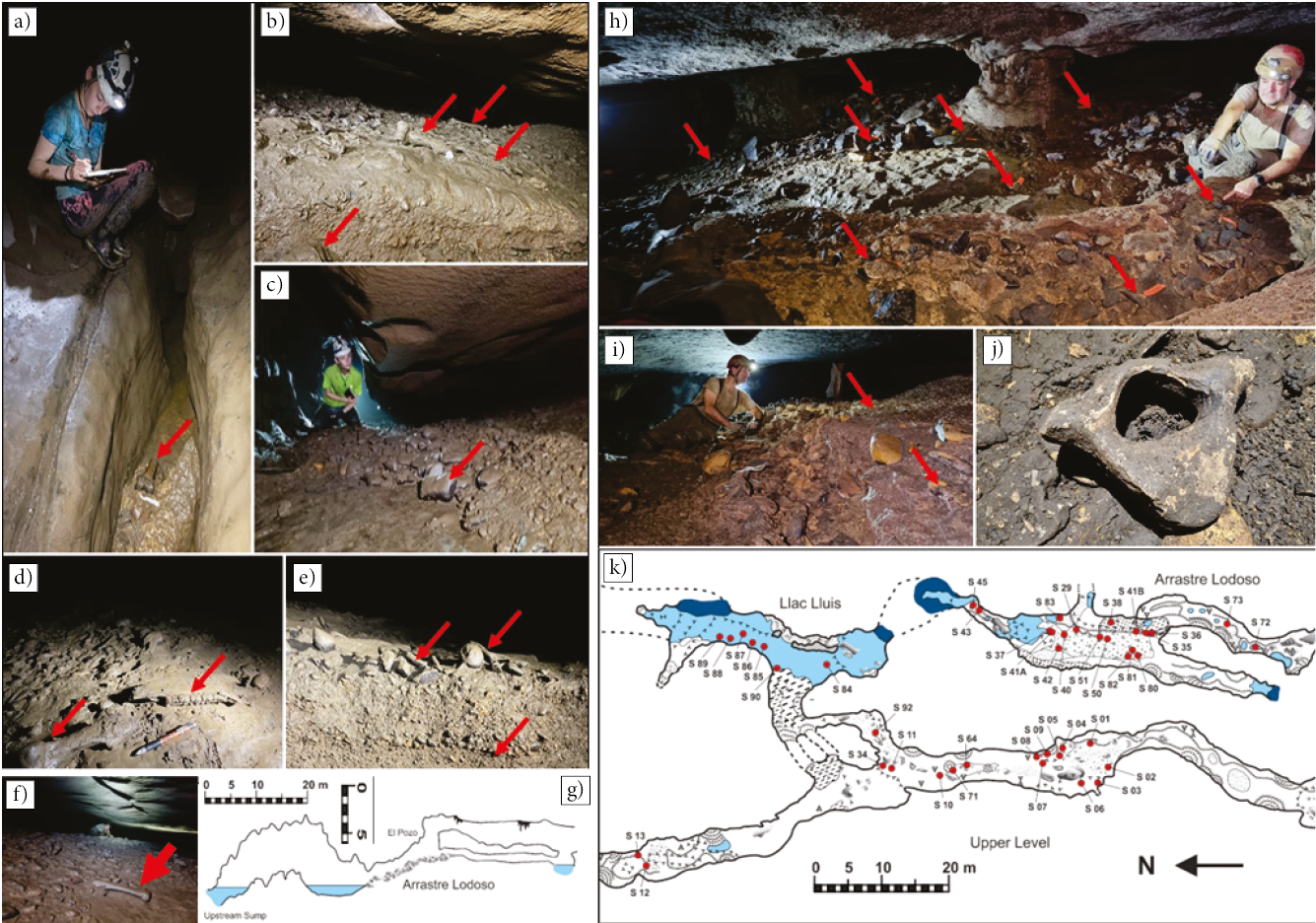
Figure 2. a-g) Views of El Arrastre Lodoso. Red arrows point at fossils. a) Sketching above a pothole containing fossils. b) Looking into El Arrastre. Fossils can be seen on top of the gravel deposit. c) Edge of the embankment of rocks and fossils. d) Fossils lying on top of the deposit and covered in still wet mud from recent flooding. e) Cross section of the deposit containing fossils, 30 cm thick. It is remobilized every rainy season. f) Inside the tight crawlway of the El Arrastre. A low ceiling is a challenge for the study of this area. One of the many bones can be seen in the forefront. g) Profile sketch of Arrastre Lodoso which explains the accumulation of fossils at the embankment, instead of being transported towards Dos Soyates. Vertical augmentation by 2X. h-k) The Upper-Level. Red arrows point at fossils. h) Overview of the passage with many fossils flagged. Notice the abundance of scattered fossils mixed with the sediments, highlighted by orange flagging throughout the gallery. i) Looking at the sediments, mostly gravel, which fills half the passage height. j) Very eroded proboscidean vertebra. k). Distribution of fossils in the middle portion of the cave.
Along the upper level, a second area of sediments and fossil concentration is around La Bifurcación, where a tunnel connects Llac Lluis to the upper level. During severe floods, water must push sediments up this tunnel and into the upper level (Figure 2k), where the sediment load, including the fossils were deposited (Figure 2h–2i).
Although in the upper level there were large specimens such as a molar of Mammuthus Brookes, 1828, and long bones and a vertebra of proboscideans, most of the fossils here were either Equus Linnaeus, 1758 teeth or very small and fragmented bones, testimony to the energy needed to lift them all the way up to this passage. The large bones show the wear and tear resulting from the transport to their resting place (Figure 2j).
Southern portion
Although Equus teeth and long bone fragments can be found throughout almost every passage in the cave, their concentration greatly diminishes outside the above-mentioned areas. Although at some point a large flood event (or events) pushed some fossils into the upper levels, most fossils are found in the lower levels. But in these passages the water flow is too strong and there are few areas where sediments (and therefore fossils) became concentrated (Supplementary Figure S9). A peccary mandible was found near the Soyate Solitario entrance but based on its lack of permineralization and color it may possibly be recent.
It should be mentioned that both prehispanic and modern human artifacts are found throughout the cave. Fragments of pottery are found in the large embankments and are particularly abundant near the entrance of Dos Soyates, where an obsidian blade of probable prehispanic origin was also found. A metal bucket, a shovel, and other metal items were seen at the pool in the Sótano del Toro 2 entrance room. Plastic bottles have been carried throughout all galleries with water flow. Sótano del Toro has a gasoline pump installed at the entrance to extract water, while Sótano del Toro 2 has also been used as a water well.
Taxonomy
This cave system preserves a large assemblage of bones with a diverse assortment of species. The faunal assemblage consists of disarticulated and mostly fragmented bones and teeth (Table 1). The number of fossils recovered to date is 93 while 16 identified specimens were left in situ. These 109 specimens are a small fraction of the total number of remains in the cave as they were surface finds, and no excavations have yet been done. Close to 300 more have been recorded in situ but await identification. Given the inherent difficulties of working within the trying conditions of a demanding cave, cataloging had to be restricted to teeth, large bones, or significant remains that were easily observed. Undoubtedly there are countless more remains that were not documented due to being small, fragmented, replicates of already recorded specimens, are under sediments, or were simply not observed when conducting general visual scanning of the surface of the sediments.
Table 1. List of identified fossils by group. Calera cave system, Sierra de El Abra.
|
Group |
Fossil type |
Total |
|
|
Artiodactyla |
Cervidae and Bovidae (Deer and Bison) |
Long bones |
2 |
|
Mandibles (with 3 teeth) |
1 |
||
|
Isolated teeth |
3 |
||
|
Antler |
1 |
||
|
Camelidae (Camels and Llamas) |
Long bones |
2 |
|
|
Mandibles (with 3 teeth) |
1 |
||
|
Isolated teeth |
1 |
||
|
Tayassuidae (Peccary) |
Mandibles (with most teeth) |
2 |
|
|
Perissodactyla |
Equus (Horse) |
Long bones |
6 |
|
Mandible (with 5 teeth) and maxilla (with 3 teeth) |
2 |
||
|
Isolated teeth |
29 |
||
|
Proboscidea |
Undetermined |
Long bones |
4 |
|
Vertebra |
1 |
||
|
Girdle |
1 |
||
|
Mammuthus (Mammoths) |
Isolated teeth |
6 |
|
|
Cuvieronius (Gomphothere) |
Isolated teeth |
1 |
|
|
Xenarthra |
Eremotherium and Megalonyx? (Sloth) |
Long bones |
2 |
|
Vertebra |
1 |
||
|
Claw |
1 |
||
|
Isolated teeth |
2 |
||
|
Carnivora |
Felidae |
Long bones |
5 |
|
Mandible (with 2 teeth) |
1 |
||
|
Isolated tooth |
1 |
||
|
Canidae: Aenocyon (Dire Wolf) |
Skull |
1 |
|
|
Testudines |
Tortoise |
Fragments of carapace |
1 |
|
Lagomorpha |
Rabbit |
Palate |
1 |
|
Undetermined |
Undetermined |
Long bones |
19 |
|
Isolated teeth |
2 |
||
|
Vertebra |
3 |
||
|
Girdle |
2 |
||
|
Rib |
4 |
||
|
Total |
109 |
||
Identification of the fossils at this time should be considered as preliminary (Table 1; Supplementary Figure S10). Future studies will help provide a more in-depth identification of the species present. From the total number of fossils identified, 54 (49.5 %) correspond to cranial and dentary elements. Of them, six were mandibles, one was a maxilla, one was a cranium, one was a palate, and 45 were individual isolated teeth. Including the 45 teeth still attached to the mandible and maxilla fragments, a total of 88 teeth were cataloged. Of the remaining fossils, 40 (36.6%) were long bones from appendages, five (4.5%) were parts of the pectoral or pelvic girdles, three (2.7%) were ribs, four (3.6%) were vertebrae, and one (0.9%) was an antler (Figure 3e). The remaining fossils have not been given an identification.
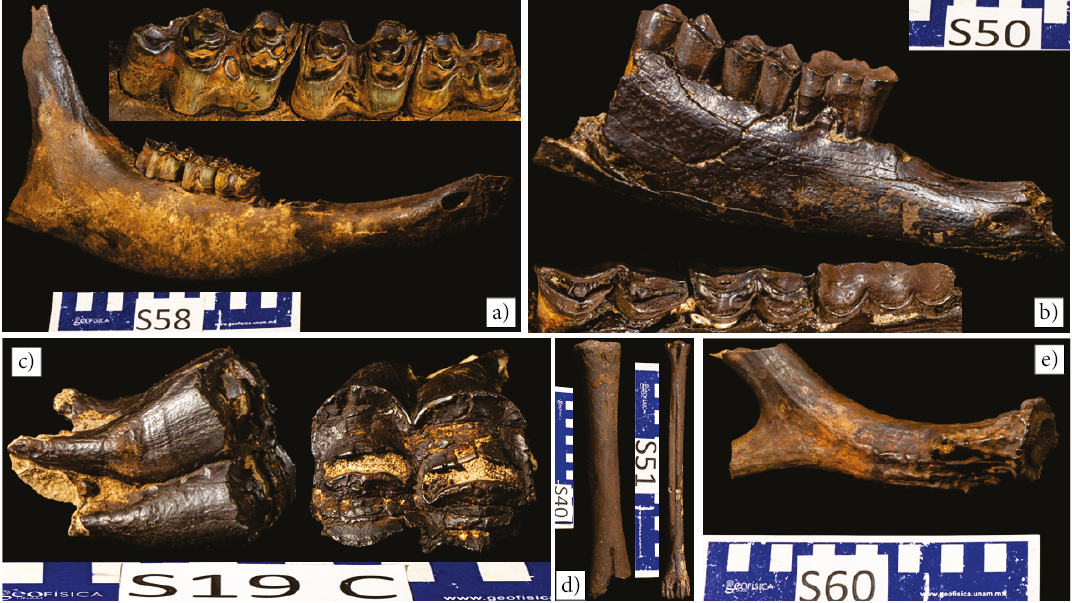
Figure 3. Artiodactyla (Camelids and Ruminants). a) Mandible of a probable Bison. b) Mandible from a juvenile camelid, the trilobed tooth at the anterior of the tooth row is the deciduous fourth premolar. c) Tooth from what may be a third species of artiodactyla. d) Metacarpal bone (left) from a camelid and metatarsal (right) from a ruminant (probably from an antilocaprid, most likely either Capromeryx or Stockoceros). e) Antler from a ruminant (deer).
Of the total number of recorded fossils (109), a preliminary taxonomic identification could be given to 79 of them (Table 1; Supplementary Figure S10b). The most common ones (37) belong to the order Perissodactyla and the dental morphology indicates they are from Equus (Figure 4). They were followed (13) by the Artiodactyla (Figure 3) and included tentatively assigned bovids based on one mandible (Figure 3a) and cervids, based on three teeth (Figure 3c), two long bones (Figure 3d right) and an antler (Figure 3e). There was also one mandible (Figure 3b), one tooth, and two leg bones (Figure 3d left) assigned to the Camelidae, and two mandibles were assigned to the Tayassuidae (Peccary; Figure 5c).

Figure 4. Perissodactyla (equids). a) Mandible. b) Fragment of Maxilla. c) Set of teeth from premolar #2 to molar #3 from the mandible of figure a). d) Canine tooth. e) Maxillary tooth showing long and curved root. f) Set of teeth from the maxilla of figure b). g) Maxillary molar #3.
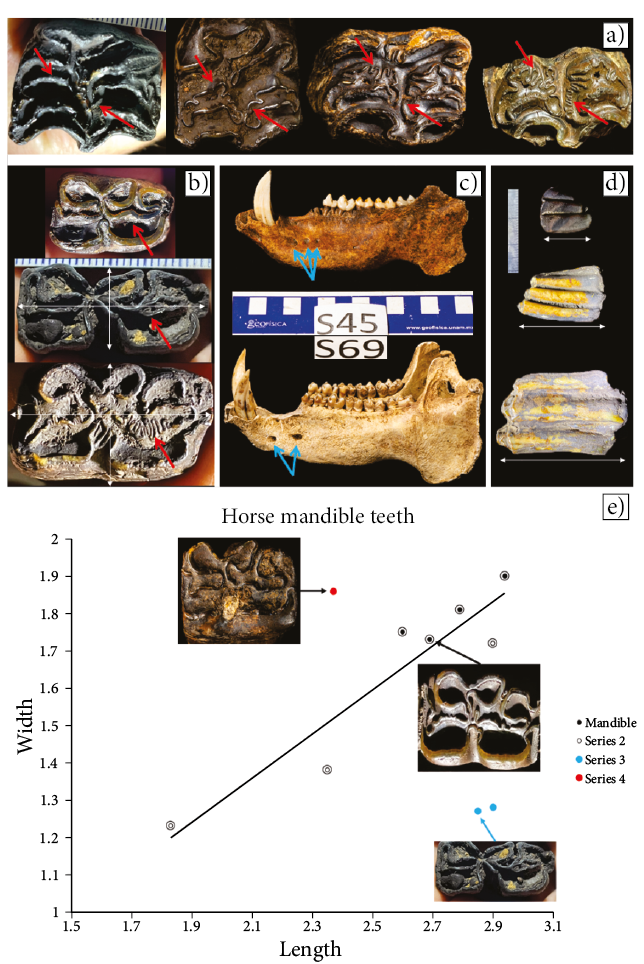
Figure 5. a-d) Diversity within population. Red arrows highlight diversity of pattern of enamel folds. White arrows highlight diversity of size. Blue arrows highlight diversity in the number of mental foramina. a) Maxilla teeth of Perissodactyla (equids). Notice diversity in enamel folds from simple (left) to complex (right). b) Mandible teeth of Perissodactyla (equids). Notice diversity in enamel folds and in size. While some teeth were short, sub-square in proportions (top and bottom), others were long, rectangular in proportions (middle). c) Mandible of Tayassuidae (peccary). Notice the different number of mental foramina and degree of fossilization. At least the lower one may be from a modern peccary. d) Middle tooth lower left third molar of an antilocaprid, probably either Capromeryx or Stockoceros. Notice differences in size. e) Diversity in size proportions of mandible teeth of Perissodactyla (equids). In solid black are the premolar and molar teeth excluding M3 from the single individual mandible S39B (Figure 2a), which show the length/width variability within an individual. All the remainder are single teeth presumably from different individuals. Three main groups can be identified; In one group (red), teeth are sub-square in shape. In another (blue), they are long and rectangular. The majority (black) fit an intermediate proportion (trendline).
Following in abundance after the Artiodactyla, there were specimens (12) of Proboscidea. Based on dentition, six could be assigned to Mammuthus (mammoth; Figure 6a–6b) and one to the Gomphothere, Cuvieronius Osborn, 1923 (Figure 6c).

Figure 6. Proboscidea (Mammoths and gomphothere). a) Lateral views of mammoth teeth. b) Occlusal views of mammoth teeth. White arrows highlight variability in size and pattern of enamel folds. c) Lateral view of Gomphothere tooth of genus Cuvieronius.
The Carnivora is represented by a fragment of a mandible (Figure 7), an isolated canine tooth, and five long bones of the family Felidae, and a canid, the cranium of a dire wolf Aenocyon Merriam, 1918 (Figure 8a).
The Xenarthra Pilosa (sloth) is represented by two long bones (Figure 9a) from Nothrotheriops Hoffstetter, 1954 band Megalonyx Jefferson, 1799, two vertebrae (Figure 9e) from Nothrotheriops, one claw from Eremotherium Spillmann, 1948 (Figure 8e), and two teeth (Figure 8d), presumably one from Eremotherium and the other from Megalonyx. Thus, there appears to be at least three species of giant sloths included in the deposits of this cave. Other Xenarthra includes Cingulata (glyptodonts), represented by fragments of the carapaces of Glyptotherium Osborn, 1903 (Figure 8b). Finally, there is also a palate from a lagomorph (Figure 8f) and fragments of carapaces of tortoise (Figure 8c).
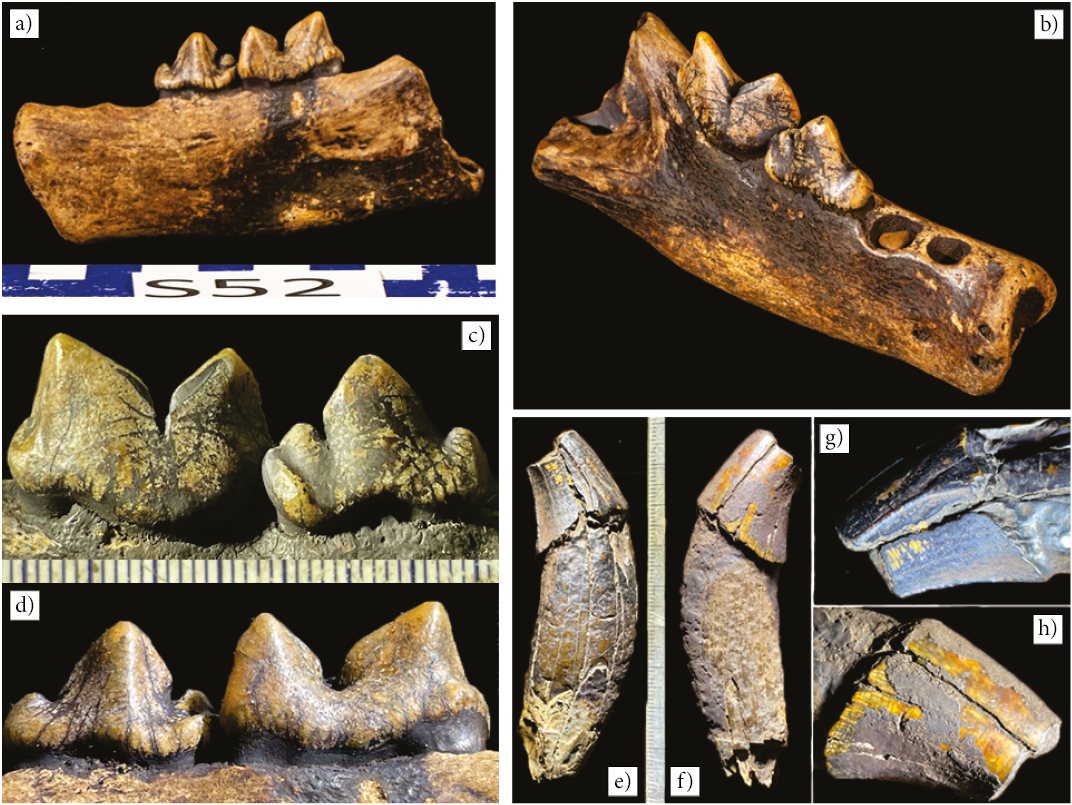
Figure 7. Felidae. a-b) Fragment of mandible. c-d) First molar (carnassial) and premolar. e-f) Canine tooth. g-h) Apex of canine tooth.

Figure 8. a) Skull of a dire wolf (Aenocyon sp.). b) Fragments of the carapace of glyptodon. c) Fragments of carapace of tortoise. d) Teeth of sloth. e) Claw of giant sloth (Eremotherium sp.). f) Palate of a lagomorph (rabbit).

Figure 9. a) Femur of a sloth with scratch marks on the edge of a bone. Squares mark areas amplified. b) Three bite and/or scratch marks in another long bone. c) Examples of the many long bones found with the shaft broken. The accumulation of such broken bones could be due to the natural mechanical stress during deposition in the cave, predation or scavenging by animals, or human breakage to access the bone marrow. d) Examples of ribs found in the cave. e) Examples of vertebrae from a sloth. f) Examples of pectoral girdle (scapula).
Morphospecies diversity
Fossils from this cave are quite abundant and, in most cases, represented by multiple skeletal elements. There are many replicates of some bones indicating multiple individuals of some species. Ongoing work in the cave will result in a larger sample size and allow for a better understanding of the diversity within groups in future studies. Variation in the large sample of teeth of the equids is probably the result of different ontogenetic stages and different degrees of abrasion of the occlusal surface during transport. Preliminary analyses suggest that within the equid group, there are some teeth that are long and thin, while there are others that are squarer in shape (Figures 5b and 5e). Likewise, the complexity of enamel folds is different among specimens (Figure 9a). Future studies will resolve if more than one species of Equus is present within the remains in this cave or if Harringtonhippus (Heintzman et al., 2017) is present.
Within the artiodactyls, the recovered dentition suggests that there may be at least three species of Camelids and Ruminants (Figure 3). As previously noted, among the proboscidean there is both mammoth and gomphothere remains (Figure 6). Within the mammoths there is a large array of premolar and molar diversity (Figure 6a–6b), suggesting that specimens of many different ontogenetic stages are represented among the remains in this cave.
Pre-burial processes
We noticed several marks on the surface of some bones that imply modification before fossilization occurred. These marks have an impregnation of manganese-oxide, suggesting that what caused the marks occurred soon after death and occurred on fresh bone. Fossil S65 (Figure 9b) has what appear to be bite marks. The presence of large felids and canids (Table 1; Figures 7 and 8a) in the fossil record of Calera system suggests that some of the faunal remains could be the result of predator hunting and/or scavenging.
Two long bones (S35 and S51; Figure 9a) and a vertebra (S05; Figure 9e) have thin and long scratching marks, which sometimes crisscross in diagnostic long crosses. It was also noticed that despite the presence of delicate, long bones such as ribs, most of the thick and sturdy bones of the appendages such as the femur or the humerus are broken (Figure 9c). It remains to be determined if they were broken prior to deposition for extracting the marrow, and if the scratch marks represent evidence of butchering by humans.
Post-burial processes
Most fossils are coated and partly permineralized by manganese oxide, as are many of the gravels that form the sediments where the fossils are emplaced (Figure 1e). Some of the fossils also have calcite encrustations on them (Figure 1h). In general, manganese oxide coated the whole surface of the pieces (Figure 1f–1g). The uniformity of the primary coatings suggests that they were formed once the bones were in the cave, under conditions of water saturation. Given that the specimens’ fossilization under water promotes the leaching out of organic carbon from the samples, we have not yet attempted radiocarbon dating of the specimen.
Cave manganese may derive from circulating water or from minor amounts of manganese in the surrounding limestone bedrock (Hill, 1982). Manganese is mobilized at the same time the limestone is dissolved and is transported in the water that percolates through the sediments in the cave (Karkanas et al., 2000). Manganese is dissolved by groundwater in the presence of CO2 by formation of the soluble hydrogencarbonate Mn(HCO3)2 (Hill, 1982). Carbon dioxide loss, oxidation, and evaporation in the cave environment concentrate the manganese in incoming solutions, whereupon the manganese precipitates forming crusts and coatings (White, 1976). Lopez-González et al. (2006) used manganese coatings to infer the bone depositional sequence in caves, but the coatings in their study are rarely as complete as those in Calera system.
All the fossils found in the upper level had an almost complete manganese-oxide coatings that usually permineralized the whole bone or tooth and many of them also had calcite coatings, particularly if they were partially buried in the sediment (Figure 1h). The fossils in the lower level also had a manganese-oxide coating, but in some cases, it was not as complete, and was restricted to the outer cortical bone and did not penetrate the whole bone, as can be observed when the fossil has broken in modern times (Figure 1f). It was also not as readily noticeable since the fossils and gravel were all covered in varying amounts of mud.
DISCUSSIONS AND CONCLUSIONS
Of the previously known Pleistocene paleontological sites, Cueva del Abra, in Tamaulipas, is the single locality for which a Pleistocene fauna has been previously described for the same mountain range as Calera system (Supplementary Figure S1). Nevertheless, despite mentioning the presence of larger bones of probable Pleistocene age, the previously published studies (Dalquest & Roth, 1970; Holman, 1970) concentrated only on smaller fossils, mostly of small mammals, rodents, bats, and herptiles with no megafauna reported.
The faunal assemblage found in Calera includes animals such as mammoths, camelids, and equids, which would prefer open grasslands. But there are also gomphotheres and cervids who preferred more forested areas. Today the Sierra de El Abra and nearby areas show a large diversity of flora, which changes with elevation, with tropical dry forest in the hills and grasslands in the coastal plain and the flat valleys of the Tantoán and Valles rivers. It is likely that during the Pleistocene times there were equally diverse environments nearby. Today the entrances to the cave are in the Tropical Dry Forest vegetation zone.
The Calera system is located at the contact between the Sierra Madre Oriental and the coastal plain (Ferrusquía-Villafrancca et al., 2017). In the two nearest sites on the coastal plain, proboscidean fossils were found; a Mammuthus molar in Xicotencatl (Riva-Palacio, 1969) and a gomphothere (Gomphotherium) in Vichinchijol (Tapia-Ramírez et al., 2013; Polaco & Aguilar, 1999). Other sites in San Luis Potosí (e.g., El Cedral, Mina San Antonio, Cueva la Presita) are located on either the Central Plateau or the Sierra Madre Oriental (Ferrusquía-Villafranca et al., 2017). The nearest one, Manantial de la Media Luna, is a karstic spring where Pleistocene megafaunal remains were also recovered (Hernández-Junquera, 1977), including Mammuthus, Equus, Camelidae and Cervidae. The similarity in types of fauna with that of Calera cave system, near the coastal plain, and Media Luna, near the Central Plateau, would seem to indicate that the Sierra Madre was not a barrier for the fauna.
Currently, it is unclear if all the fossils correspond to a single depositional event or if they accumulated from multitude events, including modern times. Near the entrance of the cave there is a small pig farm, so it is possible that people have discarded animal carcasses by disposing them into the cave. As part of the assemblage of fossils there are two mandibles of members of the suborder Suina determined to be of the Family Tayassuidae (Peccaries) and not to the Family Suidae (domestic pigs). They also happen to be the specimens with the least degree of permineralization or coating with manganese oxide. As mentioned before, the reduced (Figure 5c: S45) or lack (Figure 5c: S69) of a manganese coating on the peccary mandible leads us to believe they may be of more recent origin, indicating that bone accumulation has been an ongoing process, and the bone assemblage is most likely a time-averaged sample that potentially represents thousands of years to form. Unlike these two specimens, all the remaining fossils are heavily permineralized and, since the remains include extinct animals such as mammoths, we interpret that most are from older times.
Every time the cave floods, the fossils in the lowest active level are remobilized as part of the sedimentary load of the stream. During at least one particularly strong flood event, enough water was flushed into the system to completely flood the cave and push some of the fossils into the upper level, including fossil S16 (Figure 6a-b), the largest Mammuthus molar found. The distribution of the fossils shows that they probably entered the cave through Sótano del Toro and Sótano del Toro 2, although the second has a small entrance (40 cm wide). When trying to explain what could cause such a large and diverse megafaunal assemblage within the confines of this cave, it is obvious that some type of concentrating process occurred. We formulate three hypotheses. Future work could determine which of the three or combination of them took place.
The first hypothesis is that Sótano del Toro worked as a death trap pit. Being a karstic area, there are no surface streams nearby and the pool at the bottom of this doline is the only source of water in kilometers. The entrance has the shape of a funnel, whose edges progressively get steeper and narrower (Supplementary Figure S6a). The north side of the doline is vertical or overhanging, allowing animals to fall in and die. The south side of the doline starts with a gentle slope that could easily be walked by and would tempt animals attracted by water (Supplementary Figure S6b). But this gentle slope progressively gets steeper and steeper, with progressively bigger vertical jumps of up to 1-2 meters (Supplementary Figure S6c). It also gets narrower. An animal attracted to the water such as a mammoth could find itself unable to turn around and back off. Water during the dry season is nowadays at the bottom of a slightly overhanging 1 m wall (Supplementary Figure S6d) where, if an animal fell into the pool, it would have great difficulty escaping this trap and would drown.
The second hypothesis is that predators concentrated their kills around the waterhole. Some large felines place and stash their kills on trees. In limestone areas, large trees are preferentially found in cave entrances since it is where their roots can find water. As supporting evidence, we have the assumed bite or gnaw marks on fossil S65 (Figure 9b). Likewise, as part of the assemblage of fossils there are eight bones assigned to large carnivores (Figures 7 and 8a). There are proportionally fewer predators than prey in most environments, thus making it difficult to find large predators in the fossil record. Close to 7.3% of the identified fossils to date are from carnivore taxon, suggesting that these predators may have been abundant at this locality.
A third hypothesis is that animals were dismembered by humans, and the remains were thrown into either Sótano del Toro or Sótano Toro 2. As evidence, there are possible scratch marks in fossils S05 and S35 (Figure 9a), and the fact that most long bones found were fragments, possibly broken to extract the marrow (Figure 9c). This intriguing hypothesis will require corroboration by specialists that can unambiguously determine the origin of the scratch marks and is currently only presented as a hypothesis.
Our observations show that some of the largest remains are below the entrance pit of Sótano Toro 2. Full mandibles of equids and artiodactyls were found by snorkeling in this pool. But the 50 cm wide narrow entrance to Sótano del Toro 2 would have prevented the remains from entering the cave through that particular pit. It is suggested that the sump pool connecting Sótano del Toro with Sótano Toro 2 probably still hosts the largest bones and bone fragments that have not yet been remobilized along the cave passages downstream. Future studies involving remotely operated vehicles (ROV) or expert cave-divers may further enlarge the assembly of fossil remains discovered until now.
Acknowledgments
Jordi Espinasa for proof-reading. Marist College, Scurion, Pigeon Mountain Industries (PMI), and Stenlight for partially supporting field work.
REFERENCES
Aguayo-Camargo, J. E. (1998). The middle Cretaceous El Abra Limestone at its type locality (facies, diagenesis and oil emplacement), east-central Mexico. Revista Mexicana de Ciencias Geológicas, 15(1), 1–8.
Alberdi, M. T., Arroyo-Cabrales, J., & Polaco, O. J. (2003) ¿Cuántas especies de caballo hubo en una sola localidad del Pleistoceno Mexicano?. Revista Española de Paleontología, 18(2), 205–212, ISSN 0213-6937, http://dx.doi.org/10.7203/sjp.18.2.21648.
Alberdi, M. T., Arroyo-Cabrales, J., Marín-Leyva, A. H., & Polaco, O. J. (2014). Study of Cedral Horses and their place in the Mexican Quaternary. Revista Mexicana de Ciencias Geológicas, 31(2), 221–237.
Álvarez, R. (1977). A Pleistocene avifauna from Jalisco. Contributions of the Museum of Paleontology University of Michigan, 24(19), 205–220.
Álvarez, T., Polaco, O. J. (1982). Restos pleistocénicos de dos especies de Microtus ( Rodentia: Muridae), del norte de San Luis Potosí, México. Anales de la Escuela Nacional de Ciencias Biológicas, 26, 47–53.
Arellano, A. R. V. (1951). Research on the Continental Neogene of Mexico. American Journal of Science, 249(8), 604–616. http://dx.doi.org/10.2475/ajs.249.8.604.
Arroyo-Cabrales, J., & Polaco, O. J. (2003). Caves and the Pleistocene Vertebrate Paleontology of México. In B. W. Schubert, J. I. Mead, & R. W. Graham (Eds.), Ice age cave faunas of North America (pp. 273–291). University Press and Denver Museum of Nature & Science.
Arroyo-Cabrales, J., Polaco, O. J., Laurito, C., Johnson, E., Alberdi, M. T., & Valerio Zamora, A. L. (2007). The proboscideans (Mammalia) from Mesoamerica: Quaternary International, 169, 17–23, http://dx.doi.org/10.1016/j.quaint.2006.12.017.
Bonet, F. (1952). La facies urgoniana del Cretácico Medio de la región de Tampico. Boletín Asociación Mexicana Geólogos Petroleros, 4(5-6), 153–262.
Bonet, F. (1953). Datos sobre las cavernas y otros fenómenos erosivos de las calizas de la Sierra de El Abra. Congreso Cientifico Mexicano Memorias, 5, 238–273.
Bonet, F. (1963). Biostratigraphic notes on the Cretaceous of eastern Mexico. Corpus Christi Geological Society Annual Field Trip Guidebook, Peregrina Canyon and Sierra de El Abra, Mexico, 36-48.
Bonilla Díaz, C. (2018). Paleoecología del Pleistoceno tardío en la Cueva La Presita (San Luis Potosí, México) [Bachelor Thesis], Benemérita Universidad Autónoma de Puebla.
Boyd, D. R. (1963). Geology of the Golden Lane trend and related fields of the Tampico embayment. Corpus Christi Geological Society Field Trip Guidebook, Geology of Peregrina Canyon and Sierra de El Abra, Mexico, 49–56.
Carrillo-Bravo, J. (1971). La plataforma Valles-San Luis Potosí: Boletín Asociación Mexicana de Geólogos Petroleros, 23(1-6), 1–113.
Dalquest, W. W., & Roth, E., 1970, Late Pleistocene Mammals from a Cave in Tamaulipas, Mexico. The Southwestern Naturalist, 15(2), 217–230. http://dx.doi.org/10.2307/3670350
Dasher, G. (1994). On Station, a complete handbook for Surveying and Mapping Caves. National Speleological Society, Inc.
Day, A. (2002). Cave Surveying. British Cave Research Association, Cave Studies Series 11.
De Anda, P. (2009). La fauna local Mina de San Antonio, Pleistoceno de San Luis Potosí: Taxonomía, comparación actualística y significación geológico-paleontológica [Masters Thesis]. Posgrado en Ciencias Biológicas, Instituto de Geología, Universidad Nacional Autónoma de México.
Ellis, B. (1976). Surveying Caves. British Cave Research Association, Great Britain.
Elliott, W. R. (2015). Chapter 1. Cave exploration and mapping in the Sierra de El Abra Region. In A.C.,Keene, M. Yoshizawa, & S. E. McGaugh (Eds.), Biology and Evolution of the Mexican Cavefish (pp. 9–40). Academic Press (Elsevier). http://dx.doi.org/10.1016/B978-0-12-802148-4.00001-3
Elliott, W. R. (2018) The Astyanax Caves of Mexico, Cavefishes of Tamaulipas, San Luis Potosí, and Guerrero. Association for Mexican Cave Studies Bulletin, 26, 325 pp.
Enos, P. (1974) Reefs, platforms and basins of middle Cretaceous in northeast Mexico. American Association of Petroleum Geologists Bulletin, 58, 800–809.
Espinasa, L., & Espinasa, M. (2015). Hydrogeology of caves in the Sierra de El Abra Region. Chapter 2. In A. C. Keene, M. Yoshizawa, & S.E. McGaugh (Eds.), Biology and Evolution of the Mexican Cavefish (pp. 41–58). Academic Press (Elsevier), http://dx.doi.org/10.1016/B978-0-12-802148-4.00002-5
Espinasa, L., Ornelas-García, C. P., Legendre, L., Rétaux, S., Best, A., Gamboa-Miranda, R., Espinosa-Pérez, H., & Sprouse, P. (2020). Discovery of Two New Astyanax Cavefish Localities Leads to Further Understanding of the Species Biogeography. Diversity, 12(368), 1–21. https://doi.org/10.3390/d12100368
Ferrusquía-Villafranca, I., Ruiz-González, J. E., Torres-Hernández, J. R., Martínez-Hernández, E., & Gama-Castro, J. (2016). A new Miocene Formation from The Peotillos-Tolentino Graben fill, Western Sierra Madre Oriental at San Luis Potosí, México: Part 1, Geology. Boletín de la Sociedad Geológica Mexicana, 68(2), 247–282. http://dx.doi.org/10.18268/BSGM2016v68n2a6
Ferrusquía-Villafranca, I., Arroyo-Cabrales, J., Johnson, E., Ruiz-González, J., Martínez-Hernández, E., Gama-Castro, J., de Anda-Hurtado, P., & Polaco, O. J. (2017). Quaternary Mammals, People, and Climate Change: A View from Southern North America. In G. G. Monks, (Ed.), Climate Change and Human Responses: A Zooarchaeological Perspective, Vertebrate Paleobiology and Paleoanthropology (pp. 27–67). New York, Springer, . DOI 10.1007/978-94-024-1106-5_3
Fish, J. (2004). Karst hydrology of the Sierra de El Abra, México: Association for Mexican Cave Studies Bulletin, 14, 186 pp.
Ford, D. C. (2000). Deep phreatic caves and groundwater systems of the Sierra de El Abra, Mexico. In A.B. Klimchouk, D. C. Ford, A. N. Palmer, & W. Dreybrodt (Eds.) Speleogenesis: Evolution of Karst Aquifers (pp. 235–331). National Speleological Society.
Freudenberg, W. (1910). Die Säugetierfauna des Pliocäns u Post-Pliocäns von Mexiko. 1. Carnivoren. Geologische und Palaeontologische Abhandlungen, NS 9, 195–231.
Freudenberg, W. (1921). Geologie von Mexiko. Borntraeger.
Heintzman, P. D., Zazula, G. D., MacPhee, R. D. E, Scott, E., Cahill, J. A., McHorse, B. K., Kapp, J. D., Stiller, M., Wooller, M. J., Orlando, L., Southon, J., Froese, D. G., Shapiro B. (2017). A new genus of horse from Pleistocene North America. eLife, 6, e29944. https://doi.org/10.7554/eLife.29944
Hernández Junquera, A. (1977). Fauna local Laguna de la Media Luna, Pleistoceno Tardío, Municipio de Río Verde, San Luis Potosí, México [Bachelor Thesis]. Universidad Nacional Autónoma de México.
Hill, C. A. (1982). Origin of black deposits in caves. National Speleological Society Bulletin, 44, 15–19.
Hoffstetter, R. (1954). Les gravigrades (Edentés Xénarthres) des cavernes de Lagoa Santa (Minas Gerais, Brésil). Annales des Sciences Naturelle (Zoologie), 16, 741–764.
Holman A.J. (1970). A small Pleistocene herpetofauna from Tamaulipas. Quaterly Journal of the Florida Academy of Sciences, 32(2), 153–158.
Jefferson, T. (1799). A memoir on the discovery of certain bones of a quadruped of the clawed kind in the western parts of Virginia. Transactions of the American Philosophical Society, 4, 246–260.
Karkanas, P., Bar-Yosef, O., Goldberg, P., & Weiner, S. (2000). Diagenesis in prehistoric caves: the Use of Minerals that form in situ to assess the completeness of the archaeological record. Journal of Archaeological Science, 27, 915–929. http://dx.doi.org/10.1006/jasc.1999.0506
Keene, A.C., Yoshizawa, M., & McGaugh, S.E. (2015). Biology and Evolution of the Mexican Cavefish. Amsterdam: Academic Press (Elsevier), 412 pp.
López-González, F., Grandal-d'Anglade, A., & Vidal-Romaní, J. R (2006). Deciphering bone depositional sequences in caves through the study of manganese coating. Journal of Archaeological Science, 33, 707–717. https://doi.org/10.1016/j.jas.2005.10.006
Lorenzo L.J., & Mirambell L. (1981, Oct. 19-24,). El Cedral. S.L.P. México: Un sitio con presencia humana de más de 30000 años. Congreso de la Unión Internacional de Ciencias Prehistóricas y Protohistóricas, México, D.F. 1981, Comisón XII: el Poblamiento de América. Coloquio: Evidencia arqueológica de ocupación humana en América anterior a 11 500 años a.p. 112–124.
McDonald, H. G., Arroyo-Cabrales, J., Alarcón-Durán, I., & Espinosa-Martínez, D. V. (2020). First record of Meizonyx salvadorensis (Mammalia: Xenarthra: Pilosa) from the late Pleistocene of Mexico and its evolutionary implications. Journal of Systematic Palaeontology, 18(22), 1829–1851. http://dx.doi.org/10.1080/14772019.2020.1842816.
Marín-Leyva, A. H., Pérez-Crespo, V. A., & Torres-Hernández, J. R. (2018). Nuevo registro de los caballos del Pleistoceno Equus conversidens y E. mexicanus en San Luis Potosí, México. Paleontología Mexicana, 7(2), 81–92.
Merriam, John C. (1918). Notes on the systematic position of the wolves of the canis dirus group. Bulletin of the Department of Geology, 10(27), 531-533. https://www.biodiversitylibrary.org/item/139018
Mitchell, R. W., Russell, W. H., & Elliot, W. R. (1977). Mexican Eyeless Characin Fishes, Genus Astyanax: Environment, Distribution, and Evolution (12). The Museum, Texas Tech University Special Publications.
Osborn, H. F. (1903). Glyptotherium texanum, a new glyptodont from the lower Pleistocene of Texas. Bulletin of the American Museum of Natural History, 19, 491–494.
Osborn, H. F. (1923). New subfamily, generic, and specific stages in the evolution of the Proboscidea. American Museum Novitates, 99. http://hdl.handle.net/2246/3264
Pérez-Crespo, V. A., Arroyo-Cabrales, J., Alva-Valdivia, L. M., Morales-Puente, P., & Cienfuegos-Alvarado, E. (2012). Datos isotópicos (δ13C, δ18O) de la fauna pleistocénica de la Laguna de las Cruces, San Luis Potosí, México. Revista Mexicana de Ciencias Geológicas, 29, 299–307.
Perez-Crespo, V. A., Arroyo-Cabrales, J., Benammi, M., Polaco, O. J., Santos-Moreno, A., Morales-Puente, P., Cienfuegos-Alvarado, E., & Otero, F. J. (2013). Variación de la dieta del mamut de las praderas de Laguna de las Cruces, San Luis Potosí, México. Boletín de la Sociedad Geológica Mexicana, 65(3), 573–580.
Pichardo Del Barrio, M. (1960). Proboscídeos fósiles de México una revisión. Instituto Nacional de Antropología e Historia, Serie Investigaciones, 4, 1–63.
Polaco-Ramos, O. (1981). Restos fósiles de Glossotherium y Eremotherium Edentata en México. Anais II Congreso Latinoamericano Paleontoloia, l Porto Alegre Brasil, 820.
Polaco, O. J., & Aguilar, F. J. (1999). Fauna fósil procedente de Vichinchijol, Ébano San Luis Potosí, orden 9-99. Laboratorio de Paleozoología, Subdirección de Laboratorios y Apoyo Académico, Instituto Nacional de Antropología e Historia, México D.F., México, Informe de trabajo Z-512, 15 pp.
Polaco, O., & Butrón, L. (1997). Mamíferos pleistocénicos de la cueva La Presita, San Luis Potosí, México. In J. Arroyo-Cabrales & O. Polaco (Eds.), Homenaje al Dr. Ticul Álvarez (pp. 279–296). México D.F.: Instituto Nacional de Antropología e Historia, Colección Científica.
Reynoso, V. H., & Montellano-Ballesteros, M. (2004). A new giant turtle of the genus Gopherus (Chelonia: Testudinidae) from the Pleistocene of Tamaulipas, México, and a review of the phylogeny and biogeography of gopher tortoises. Journal of Vertebrate Paleontology, 24(4), 822–837. DOI:10.1671/0272-4634(2004)024[0822:ANGTOT]2.0.CO;2
Riva-Palacio, E. J. (1969). Megafósiles de la Planicie del Guayalejo, Tamaulipas. Boletín de la Asociación Mexicana de Geólogos Petroleros, 2(1-6), 26–35.
Russell, W.H., & Raines, T. (1967). Caves of the Inter-American Highway, Nuevo Laredo, Tamaulipas to Tamazunchale, San Luis Potosí. Association for Mexican Cave Studies Bulletin, 1, 126 pp.
Silva-Barcenas, A. (1969). Localidades de vertebrados fósiles en la República Mexicana. Palentología Mexicana, 28, 1–52.
Spillmann, F. (1948), Beiträge zur Kenntnis eines neuen gravigraden Riesensteppentieres (Eremotherium carolinense, gen. et sp. nov.) seines Lebensraumes und seiner Lebensweise. Palaeobiologica, 8, 231–279.
Tapia-Ramírez, G., Guzmán, A. F., & Polaco, O. J. (2013). Los gonfoterios (Proboscidea, Gomphotheriidae) de Colima, México. Boletín de la Sociedad Geológica Mexicana, 65(3), 591–607.
Torres Martínez, J. C. (1995). Fauna local Mina de San Antonio, pleistoceno tardío de San Luis Potosí, y su significación geológico-paleontológico [Bachelor Thesis]. Universidad Autónoma de Morelos.
Voorhies, M. R. (1969). Taphonomy and population dynamics of an Early Pliocene Vertebrate Fauna, Knox County, Nebraska. Contributions to Geology, University Wyoming, Special Paper, 1, 69, https://doi.org/10.2113/gsrocky.8.special_paper_1.1
White, W. B. (1976). Cave minerals and speleothems,. In T. D., Ford, C.H.D. Cullingford (Eds.), The Science of Speleology (pp. 267–327). London, Academic Press.
