Revista Mexicana de Ciencias Geológicas, v. 42, núm. 1, abril 2025, p. 31–43

Historia de la vegetación y el clima durante los últimos 3200 años, en la costa sur de Quintana Roo, México
Chloe Rosas1, Alicia Carrillo-Bastos1,*, Claudia González-Salvatierra2, Alejandro Antonio Aragón-Moreno3 y Gerald Islebe3
1 Tecnológico Nacional de México / IT de Chetumal, A. Insurgentes No. 330, Esq. Andrés Q. Roo, Col. David Gustavo Gutiérrez, C.P. 77013, Chetumal, Quintana Roo, México.
2 SECIHTI, Tecnológico Nacional de México / IT de Chetumal, Av. Insurgentes No. 330, Esq. Andrés Q. Roo, Col. David Gustavo Gutiérrez, C.P. 77013, Chetumal, Quintana Roo, México.
3 El Colegio de la Frontera Sur, Unidad Chetumal, Avenida Centenario Km 5.5, C.P. 7700, Chetumal, Quintana Roo, México.
* Autor para correspondencia (A. Carrillo-Bastos): alicia.cb@chetumal.tecnm.mx
RESUMEN
Por medio de análisis de polen y elementos geoquímicos, de un núcleo de sedimento de 1.5 m de profundidad, se identificaron cambios en la historia vegetal y ambiental de los últimos 3200 años de la Laguna Cementerio ubicada en el poblado de Xcalak, Quintana Roo. Los cambios en la composición y abundancia de Rhizophora mangle, Conocarpus erectus, Avicennia germinans y Laguncularia racemosa permitieron inferir cambios en la variabilidad hidrológica. Al inicio de los 3200 años cal AP se presentaron condiciones húmedas, la vegetación dominante era la selva, compuesta principalmente por elementos de Moraceae, Fabaceae, Euphorbiaceae, Rubiaceae y Bursera simaruba. Durante este periodo, los elementos de manglar estaban dominados por Avicennia germinans junto con la presencia de foraminíferos. El primer registro de Zea mays alrededor de 850 a.C (2800 cal AP) permitió identificar la presencia de actividades agrícolas cerca de la zona desde el Preclásico temprano hasta el Clásico tardío. La especie de manglar dominante era C. erectus, lo que sugiere fuertes cambios en la variabilidad hidrológica. Se lograron identificar, por medio de la curva de Ca/Σ (Ti, Fe, Al), dos periodos de sequía alrededor de 1700 y 850 cal AP, respectivamente. La sequía alrededor de 1700 cal AP podría estar asociada a una intensificación de ENSO y a un cambio en el gradiente de la temperatura superficial del mar y la salinidad que afectan la fuerza de la corriente de chorro del Caribe, provocando una disminución en la cantidad de precipitación para Mesoamérica. Durante ese mismo periodo se registró la mayor reducción de la selva y expansión de la sabana, lo cual permitió el depósito de Pinus sp. La sequía registrada alrededor de 850 cal AP es contemporánea con el periodo Clásico terminal. La comparación con otros registros climáticos de la región y del Circum-Caribe sugieren que las sequías del Clásico terminal estaban alternadas con periodos húmedos. El regreso de las condiciones húmedas alrededor de 700 cal AP permitió la recuperación de la cobertura vegetal. El presente estudio es un registro que aporta nuevas evidencias sobre los cambios en la Laguna Cementerio durante los últimos 3200 años, zona que ha sido poco estudiada desde el punto de vista paleoambiental.
Palabras clave: vegetación; manglar; paleoambientes; polen fósil; geoquímica; sequía; Quintana Roo; México.
ABSTRACT
Changes in vegetation and environmental history for the last 3200 years at Laguna Cementerio, Xcalak, Quintana Roo were identified using fossil pollen and geochemical analyses of a 1.5 m depth sediment core. Changes in abundance and composition of Rhizophora mangle, Conocarpus erectus, Avicennia germinans, and Laguncularia racemosa allowed to infer changes in hydrological variability. At the beginning of the 3200 cal yr BP moist conditions were exhibited. Dominant vegetation was tropical forest, mainly composed of Moraceae, Fabaceae, Euphorbiaceae, Rubiaceae, and Bursera simaruba. During this period, mangroves were dominated by Avicennia germinans along with the presence of foraminifera. The first record of Z. mays around 850 B.C. (2800 cal yr BP) allowed the identification of agricultural activities close to the study area from the Middle Preclassic to the Late Classic. The dominant mangrove species was C. erectus, suggesting strong changes in the hydrological variability. Two drought periods were identified around 1700 and 850 cal yr BP, respectively, with the Ca/Σ (Ti, Fe, Al) curve. The drought at 1700 cal BP may be associated with changes in sea surface temperatures (SST) and sea surface salinity (SSS) that affect the strength of the Caribbean Low-Level Jet causing a decrease in the amount of rainfall for Mesoamerica. During this same period, the highest reduction of tropical forest was observed, changing the landscape to a more open vegetation that allowed the deposit of Pinus sp. The drought that occurred around 850 cal yr BP corresponds to the Terminal Classic Period. Further comparison with other climate proxy records from the region and Circum-Caribbean area suggests that the Terminal Classic Droughts were alternated with wet periods. Around 700 cal yr BP, moist conditions returned and allowed the recovery of the tropical forest. This study contributes to new evidence of changes experienced in Laguna Cementerio during the last 3200 cal yr BP, an area that has been poorly studied from a paleoenvironmental point of view.
Keywords: vegetation; mangrove; paleoenvironment; fossil pollen; geochemistry; drought; Quintana Roo; Mexico.
Manuscrito recibido: diciembre 16, 2024
Manuscrito corregido recibido: marzo 10, 2025
Manuscrito aceptado: marzo 24, 2025
Publicado en línea: abril 1, 2025
INTRODUCCIÓN
Los ecosistemas de manglar son sensibles al cambio climático y los cambios en el patrón regional de precipitación pueden influir en el aporte de agua dulce, en la descarga de sedimentos y nutrientes causando cambios en la estructura y distribución de estos ecosistemas (Punwong et al., 2018). Los manglares son altamente adaptativos con tolerancia a condiciones ambientales extremas tales como alta salinidad, anoxia, inundación, y temperaturas (Lovelock et al., 2015). Esta adaptabilidad ha permitido que este ecosistema resista cambios ambientales a lo largo del Holoceno (Monacii et al. 2009), donde han existido periodos variables en temperatura y precipitación (Hodell et al., 2001; Roberts, 2014). Por ejemplo, en Belice, Wooller et al. (2009) documentó que los individuos de la especie Rhizophora mangle pasaron de ser enanos a ser individuos de transición (individuos altos) como respuesta a los cambios en el nivel del mar y precipitación.
Los depósitos sedimentarios costeros son archivos importantes de cambios paleoambientales ampliamente utilizados para entender el desarrollo, adaptación, y resiliencia de los ecosistemas costeros ante cambios ambientales en el pasado, permitiendo así conocer los cambios en el nivel del mar desde una perspectiva temporal amplia (Urrego et al., 2009; Sefton & Woodroffe, 2021). En particular, los sedimentos de manglar proporcionan condiciones adecuadas para la preservación de registros orgánicos como el polen: condiciones anaeróbicas y reductivas, bajo pH, alta tasa de acumulación y una alta salinidad (Phuphumirat et al., 2015; Liu et al., 2024), y a través de su análisis se pueden evaluar los cambios en la vegetación a lo largo del tiempo (Brewer et al., 2013; Friedel et al., 2024) a fin de reconstruir la historia de la vegetación en un área determinada. La alta sensibilidad de los ecosistemas de manglar, junto con la composición florística simple (baja diversidad, alta dominancia) y el marcado gradiente de salinidad, facilita su identificación en los registros palinológicos, con lo que es posible la determinación de cambios tanto a nivel local como regional (Aragón-Moreno et al., 2012).
El inicio del Megalayiano hace 4200 años estuvo marcado por una disminución significativa en la precipitación con respecto al Norgripiano (Haug et al. 2001; Wooller et al. 2007; Mueller et al. 2009; Walker et al. 2019). En la Península de Yucatán, el Megalayiano se ha caracterizado por presentar periodos de sequía alternadoss con fases húmedas (Curtis et al., 1996; Hodell et al., 2005; Aragón-Moreno et al. 2012; 2018a). En este contexto, algunos eventos importantes asociados a las anomalías climáticas y reportados por estudios paleoecológicos en la región son el establecimiento de la línea de costa, cambios en la vegetación y la detección de sequias relacionadas al colapso de la cultura maya. (Curtis et al., 1996; Torrescano-Valle & Islebe, 2006; Hodell et al., 2005; Carrillo-Bastos et al., 2013; Aragón-Moreno et al., 2012, 2018b, 2024). La vegetación a lo largo del Megalayiano ha respondido a estos cambios climáticos y diversas investigaciones en la Península de Yucatán han demostrado que la vegetación dominante al inicio del Megalayiano era la selva tropical, resultado de un clima húmedo que prevaleció durante el Norgripiano pero que progresivamente fue cambiando como respuesta a un clima menos seco (Carrillo-Bastos et al., 2010; Gutiérrez-Ayala et al., 2012; Aragón-Moreno et al., 2012; 2018a). Después de 2800 cal AP, el clima en la región era más seco y se registraron numerosas sequías (Hodell et al., 2007; Roy et al., 2017). Así mismo, durante el Megalayiano se registran las prácticas agrícolas en la región, las cuales junto con las anomalías climáticas provocaron una presión simultánea sobre la cobertura vegetal. A nivel regional, se ha registrado que los cambios en la composición de la vegetación están asociados a cambios macroclimáticos en la precipitación (Haug et al., 2003).
Investigaciones paleoecológicas realizadas en la Península de Yucatán en ecosistemas de manglar han documentado la respuesta de los manglares ante fenómenos de distintas escalas espacio-temporales, como cambios en el régimen de precipitación asociado al desplazamiento de la Zona de Convergencia Intertropical (ITCZ por sus siglas en inglés) y el fenómeno de El Nino -Oscilación del Sur (ENSO), cambios en la fuerza de la corriente de chorro del Caribe, cambios en el gradiente de temperaturas superficiales del mar, cambios en el nivel del mar, y actividades antrópicas durante el Megalayiano (Torrescano-Valle & Islebe, 2006; Wooller et al., 2007; Monacci et al., 2009, 2011; Aragón-Moreno et al., 2018a, 2024). Así mismo, diversos estudios han demostrado que al inicio del Megalayiano, se registró una disminución en la precipitación que tuvo efecto sobre los ecosistemas de manglar como resultado de una acción conjunta del aumento de la actividad del ENSO y la migración de la ITCZ hacia el sur (Haug et al., 2001; Conroy et al., 2008; Joo-Chang et al., 2015). En la Laguna Cementerio se han estudiado otros proxies biológicos para entender las variaciones climáticas utilizando los palinomorfos no polínicos (Can-Canales et al. 2023). No obstante, en esta región ecológicamente diversa, se requiere de un entendimiento de la respuesta de los sistemas de manglar ante los cambios ambientales que se presenten por forzamientos climáticos locales o regionales para el desarrollo de estrategias de conservación de la línea de costa y los ecosistemas asociados, incluyendo los manglares. Por lo tanto, el objetivo del presente estudio fue realizar una reconstrucción de la historia de las comunidades vegetales y ambiental de la zona sur de Quintana Roo, para identificar los cambios en la vegetación y del ambiente ocurridos durante los últimos 3200 años a partir de polen fósil y elementos geoquímicos en sedimentos. Por lo anterior, se plantearon las siguientes preguntas: (i) ¿Cómo la dinámica hidrológica ha influido en la conformación actual del ecosistema de manglar en la Laguna Cementerio durante los últimos 3200 años? (ii) ¿Qué fenómenos climáticos locales/globales influyen en los cambios de precipitación? (iii) ¿Cuál ha sido la respuesta de la vegetación ante la perturbación antropogénica?
MATERIALES Y MÉTODOS
Área de estudio
La Laguna Cementerio se encuentra ubicada en el poblado de Xcalak al sur del estado de Quintana Roo, en el municipio de Othón P. Blanco y se localiza en las coordenadas 18°15'27"N y 87°50'38"O (Figura. 1). El clima del área es sub-húmedo con régimen de lluvias de verano y muy cálido, con una temperatura media anual mayor a 26 °C (Vidal-Zepeda, 2005; Bautista & Palacio, 2011), con máxima de 35 °C y mínima de 14 °C. Presenta una precipitación media anual de 1200 a 1500mm (Koppen, modificado por García, 1973). Por su localización, los vientos alisios dominan durante todo el año sobre la región, con una fuerte componente del este, predominando la dirección este-sureste. Los vientos dominantes provienen del sureste en los meses de febrero a mayo y de junio a octubre del este (Vidal-Zepeda, 2005).
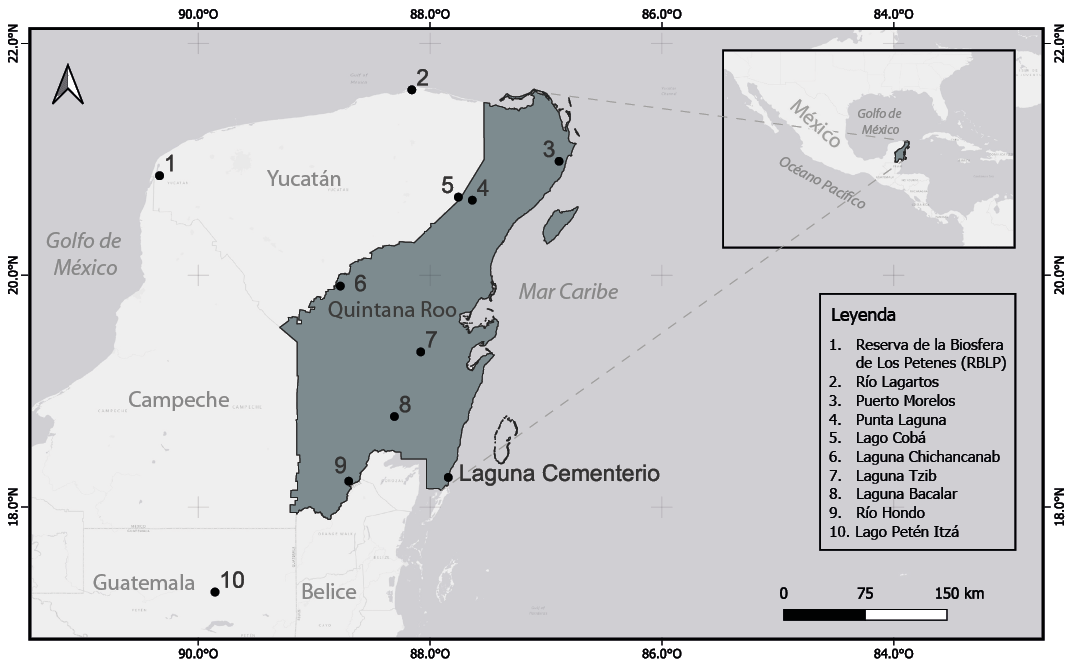
Figura 1. Área de estudio donde se indica la localización del sitio donde fue colectado el registro de sedimento.
La franja costera presenta una playa angosta y un humedal compuesto por manglares, con dominancia local de Rhizophora mangle, que se extiende a lo largo de la costa caribeña de México (Islebe et al., 2015). Dentro de la vegetación halófita en la cercanía con el manglar con tres especies dominantes (Rhizophora mangle, Avicennia germinans y Conocarpus erectus) (Comisión Nacional de Áreas Naturales Protegidas, 2004); también se presentan especies arbóreas de entre 8 y 12 m de altura como Cocos nucifera, Piscidia piscipula y Thrinax radiata, Coccoloba uvifera, Pouteria campechiana y Metopium brownei. La vegetación circundante al área de estudio está compuesta por manglar de franja de baja estatura dominado principalmente por Rhizophora mangle, 1–3 m en altura.
Respecto a la geología, Quintana Roo es parte de una estructura de origen sedimentario formada durante el Cretácico Tardío, sobre la cual se depositaron arenas y estructuras de origen orgánico marino a lo largo del Cenozoico (Álvarez, 1954; Ordoñez-Crespo & García Rodríguez, 2010). Durante el Cretácico y hasta el Eoceno tuvieron lugar una serie de transgresiones marinas que permitieron el depósito y la formación de estratos de caliza. No obstante, en el Cretácico prevalecieron las condiciones necesarias para la formación de material evaporítico que permitieron el depósito de estratos de hasta 1300 m de espesor (Ordoñez-Crespo & García-Rodríguez, 2010). Esta plataforma está formada por rocas carbonatadas, evaporíticas y clásticas tales como caliza, dolomita, yeso y arenisca (Fragoso Servón et al. 2014).
Trabajo de campo
En la Laguna Cementerio, en septiembre de 2018 se colectó un núcleo de sedimento de 150 cm de longitud, mediante un sondeador tipo ruso. Cada segmento de 50 cm fue cuidadosamente empaquetado en campo y procesado en el laboratorio de Zoología del Instituto Tecnológico de Chetumal, para su determinación cronológica, obtención de polen fósil y análisis geoquímico.
Establecimiento de la cronología
La cronología se estableció por medio de mediciones de radiocarbono AMS en la materia orgánica de tres muestras de sedimento. Las muestras fueron procesadas en el laboratorio International Chemical Analysis en Damascus, Estados Unidos de América (Tabla 1). El modelo edad/profundidad y la curva de sedimentación fueron obtenidos utilizando el paquete de software Bacon versión 4.2.3 (Blaauw & Christen, 2011), disponible en el software R (R Core Team, 2023) utilizando la curva de calibración IntCal20 (Reimer et al., 2020).
Tabla 1. Edad radiocarbono de materia orgánica para el núcleo LC.
|
Código de laboratorio |
Profundidad (cm) |
Edad 14Cal AP |
14C Error ± |
Edad media (cal AP) |
Tasa de sedimentación |
|
14C-5897 |
40.5 |
600 ± 30 |
30 |
615 |
0.0671 |
|
14C-7725 |
75 |
1840 ±30 |
30 |
1739 |
0.0301 |
|
14C-7726 |
110 |
2690 ±30 |
30 |
2789 |
0.0317 |
Polen fósil y análisis geoquímico
Se procesaron 35 muestras para el análisis polínico, obtenidas a intervalos de 4 cm. La extracción de polen se realizó mediante técnicas de extracción estándar, KOH, HCL y acetólisis (Faegri & Iversen, 1989) y montadas con medio Entellan. Se contó un mínimo de 300 granos de polen en cada muestra mediante microscopia óptica (400X), excluyendo las esporas de hongo y especies acuáticas en la suma de polen. La identificación del polen se hizo a nivel taxonómico de familia y en algunos casos, donde fue posible, a nivel de género y especie, con la ayuda de atlas palinológicos (Lozano-García & Martínez-Hernández, 1990; Palacios Chávez et al., 1991). El diagrama de polen fue graficado en el programa Tilia v3.0.3 (Grimm, 2016). Las zonas polínicas fueron definidas a través de un análisis de agrupamiento aglomerativo, jerárquico y estratigráficamente restringido por el método de suma total de cuadrados CONISS (Constrained Cluster Analysis by Sum of Square) (Grimm,1987).
Para determinar la composición elemental del sedimento, un total de 58 muestras (1 cm3 cada 2.5 cm) fueron secadas a 50 °C durante 24 horas y pulverizadas finamente con un mortero de ágata. Las muestras se analizaron con el equipo Thermoscientific Niton XL3t X-ray fluorescence (XRF) analyzer. Los datos se corrigieron usando la ecuación de regresión lineal de acuerdo con el método de Quiroz-Jiménez y Roy (2017). Sólo aquellos elementos con concentraciones por arriba de los límites de detección y que demostraron una variabilidad significativa a lo largo del registro fueron considerados en la interpretación. Los resultados fueron expresados en porcentajes y graficados.
RESULTADOS
Cronología y estratigrafía
Con el modelo edad/profundidad se estimó una edad basal de 3200 cal AP, con una tasa de sedimentación promedio de 0.043 cm/año (Figura 2). En cuanto a la estratigrafía, se observaron cinco fases de cambio en las cuales se detectaron variaciones en el contenido de materia orgánica, carbonatos y arena fina a lo largo del sedimento. El primero, entre 150–127 cm, de color café oscuro con alto contenido de materia orgánica; el segundo (70–126 cm), un bandeo de color gris claro con presencia de restos vegetales y calcáreos; el tercero (51–69 cm), se caracterizó por la presencia de restos calcáreos y baja cantidad de materia orgánica, de color gris oscuro; el cuarto (30–50 cm), color gris claro con alto contenido de arenas finas y, por último, una capa (0–29 cm) con alto contenido de materia orgánica y restos vegetales.
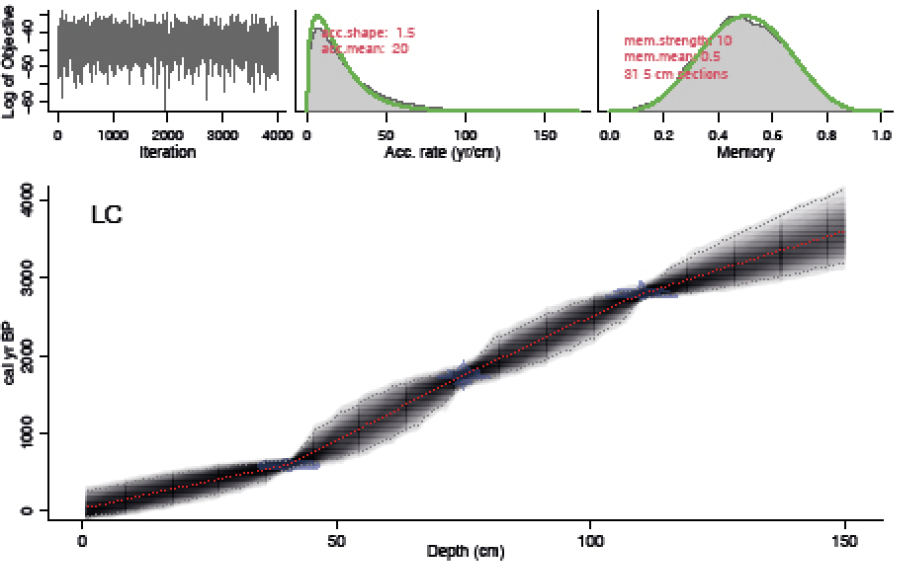
Figura 2. Modelo edad/profundidad y tasa de sedimentación (cm/año) para la Laguna Cementerio (LC). Cuadro izquierdo superior muestra las iteraciones de MCMC (Markov Chain Monte Carlo). Cuadro medio superior muestra la distribución de la tasa de acumulación (cm/año) y cuadro derecho superior muestra la memoria. Panel inferior muestra las edades 14C calibradas (azul transparente) y el modelo edad-profundidad (gris oscuro indica las edades calendario probables. Las líneas grises punteadas muestran un intervalo de confianza del 95%; línea roja muestra el mejor modelo basado en la edad media para cada profundidad).
Registro palinológico y análisis de sedimentos por XRF
En el análisis de polen fósil se identificaron un total de 52 taxas distribuidos en 42 familias. Los porcentajes para cada taxón fueron representados en el diagrama de polen, agrupados por tipo de vegetación de acuerdo con su afinidad ecológica en manglar, selva y disturbio (Figura 3). Con los cambios en la composición y a través del análisis de clúster de CONISS se delimitaron tres zonas polínicas.

Figura 3. Diagrama de polen representando los porcentajes totales de polen de manglar, selva tropical y taxa de disturbio en un sedimento recuperado de la Laguna Cementerio, en las costas sur de Quintana, Roo. Los grupos de taxa fueron agrupados de acuerdo con sus afinidades ecológicas y las zonas polínicas fueron determinadas por CONISS (Grimm, 2016).
En todas las muestras, las concentraciones de Mn, Zr, Pb y Zn se encontraron por debajo de los limites detectables del equipo (<0.01%). Sin embargo, se detectaron las concentraciones de Ti (0.0468–0.33%, promedio 0.087%), Al (0.18–3.39%, promedio 0.59%), Fe (0.13–1.05%, promedio 0.25%), Mg (1.14–2.60, promedio 1.76%), Ca (2.84–41.9%, promedio 29.4%) y K (0.17–0.80%, promedio de 0.29%) (Figura 4). El Ti se encontró en la mayor parte de las muestras, excepto en las profundidades 57, 72 y 80 cm donde los niveles estuvieron por debajo de los limites detectables del equipo. Por otro lado, las concentraciones de Ti, Al, Fe y K fueron relativamente bajas, mientras que para Ca fueron altas en comparación con los valores promedios de referencia de la Corteza Continental Superior (UCC, por sus siglas en inglés) (Taylor & McLennan, 1985), reflejando la influencia geológica de la región (Figura 4). La Península de Yucatán emergió al final del Cenozoico y es de naturaleza kárstica, estando principalmente compuesta por calcita, dolomita y yeso (Bautista et al., 2011).
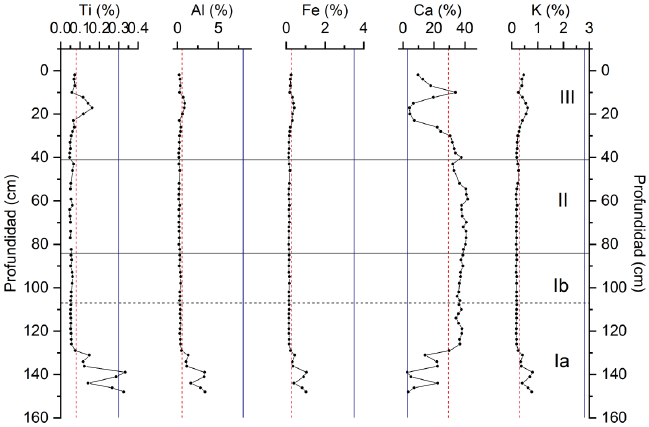
Figura 4. Concentración de Ti, Al, Fe, Ca y K a lo largo de 3200 cal AP para laguna Cementerio. Los valores promedio de cada elemento están representados por las líneas punteadas color rojo y la línea punteada azul representa el valor promedio de referencia de la Corteza Continental Superior (UCC, por sus siglas en inglés) (Taylor & McLennan, 1985).
Para reconstruir cambios pasados en la relación de E/P (Evaporación/Precipitación), se utilizó el proxy geoquímico Ca/Σ (Ti, Fe, Al); altas proporciones reflejan condiciones de sequía y bajas proporciones condiciones de humedad (Mueller et al., 2009). De acuerdo con Roy et al., (2018) la naturaleza kárstica de la Península de Yucatán tiene un efecto de dilución en las concentraciones elementales, lo que provoca bajos niveles elementales. La variación de la relación Ca/Σ (Ti, Fe, Al) en la Laguna Cementerio se asemeja al de Ca, con variaciones a lo largo del sedimento entre 0.60 y 123.25 y con un promedio de 57.9 (Figura 5). A continuación, se describen los cambios identificados en cada zona polínica junto con las características geoquímicas del sedimento.
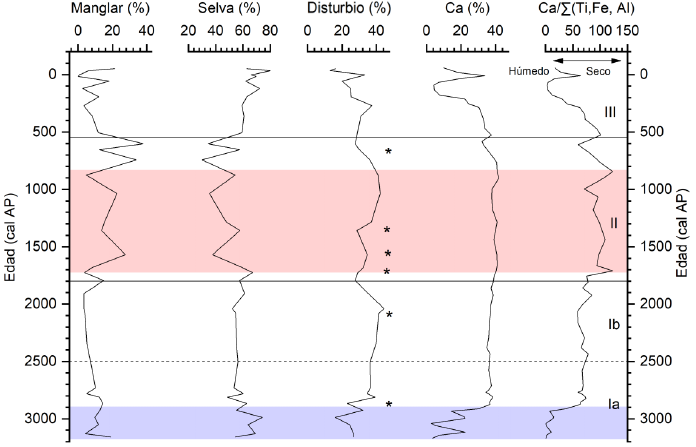
Figura 5. Porcentajes de polen correspondientes a manglar, selva, disturbio, calcita y Ca/∑ (Ti, Fe, Al) de Laguna Cementerio. La línea punteada y líneas sólidas muestran la división de zonas definida por el CONISS. La banda azul representa el periodo de aumento de humedad y la banda roja representa los periodos de mayor sequía para la Laguna Cementerio. Los asteriscos (*) representan el polen de Zea mays a lo largo del registro.
Zona I (150–84 cm; ~3200–1800 cal AP)
Está caracterizada por la presencia de altos porcentajes de selva que disminuyen a lo largo de esta zona. Esta zona está subdividida en dos subzonas, subzona Ia y subzona Ib que se describen a continuación.
Subzona Ia (150–107 cm; ~3200–2500 cal AP)
Avicennia germinans es la especie dominante del manglar, pero sus porcentajes varían ampliamente durante este periodo (5–13%). A lo largo de esta subzona se observaron foraminíferos los cuales coinciden con la presencia de Avicennia germinans en la parte inferior de esta subzona. Posteriormente, los porcentajes en los taxa de manglar fluctúan hacia valores relativamente altos hasta entre 3200–3000 cal AP (150–135 cm) cuando esta variabilidad disminuye y Conocarpus erectus es la especie dominante (5–7%), alrededor de 2850 cal AP (125 cm). En esta subzona, los taxa correspondientes a la selva están representados principalmente por Ficus (5–12%), Moraceae (7–25%), Fabaceae (14–30%) y Euphorbiaceae (3–8%); en menor proporción Bursera simaruba, Brosimum alicastrum, Rubiaceae y Combretaceae (<6%). Haematoxylum campechianum, Burseraceae, Boraginaceae y otros taxas de selva aparecen intermitentemente en abundancias <3%. Poaceae representa el 16% del polen total en esta subzona con picos de 24% alrededor de 2900 cal AP. Amaranthaceae y Asteraceae se encuentran bien representadas (1–13%) y a lo largo de toda la zona. Al final de este periodo, alrededor de 2800 cal AP (116 cm), se registra el primer grano de polen de Zea mays junto con Pinus sp. y Alnus que aparecen con valores ≤1%.
Al inicio de la subzona las concentraciones de Ti (1.66–3.39 %), Al (0.40–1.05%), Fe (0.40–1.05%) y K (0.41–0.80%) presentan porcentajes por encima del promedio, mientras que Ca (2.84–22.45%) presenta valores por debajo del promedio, con los porcentajes más bajos de todo el sondeo. Así, los porcentajes de Ti, Al, Fe, y K registrados en esta subzona fueron los porcentajes más altos del sondeo. No obstante, alrededor de 3000 cal AP (136 cm) los porcentajes de Ti, Al, Fe y K disminuyen de manera significativa y los porcentajes de Ca aumentan exponencialmente. La proporción Ca/∑ (Ti, Fe, Al) presentó un comportamiento similar al de Ca.
Subzona Ib (107–84 cm; ~2500–1800 cal AP)
Esta subzona se caracteriza por una ligera disminución de los elementos de manglar. Al inicio de este periodo Avicennia germinans es la especie dominante (1–4%), pero alrededor de 2300 cal AP (103 cm) hasta el final de esta subzona Conocarpus erectus es más abundante con valores entre 1–8%. Rhizophora mangle y Laguncularia racemosa aparecen de manera intermitente a lo largo de este periodo (1–16%). Los elementos que corresponden a selva disminuyen con respecto a la subzona Ia, en donde Fabaceae registra un aumento significativo (10–30%). Los taxa de selva como Moraceae, Ficus y Euphorbiaceae se observan a lo largo de este periodo (>15%). Poaceae continúa siendo el taxon dominante con valores máximos de 28% alrededor de 2100 cal AP (96 cm). Amaranthaceae, Asteraceae y Convolvulaceae muestran porcentajes entre 1–13% y Pinus sp. aparece de manera discontinua a lo largo de la zona. Después de 2000 cal AP desaparecen los foraminíferos. El polen de maíz se registra nuevamente al final de este periodo.
El sedimento en esta subzona presentó valores de Ca por encima del promedio (35.04–38.68%). Ti (0.05–0.06%), Al (0.27–0.40%), Fe (0.14–0.17%) y K (0.17–0.19%) se mantienen estables sin cambios significativos a lo largo de esta subzona. La proporción Ca/Σ (Ti, Fe, Al) (58.77–84.63) está por encima del promedio en esta subzona. No obstante, presenta una disminución importante en los sedimentos depositados alrededor de 2390 cal AP (104 cm), 2160 cal AP (98 cm) y 2060 cal AP (95 cm).
Zona II (84–41 cm; 1800– 550 cal AP)
La transición a esta zona se caracteriza por un aumento significativo en los elementos de manglar, al inicio de la zona Conocarpus erectus es el taxon dominante con valores ≥11%. A partir de 1000 cal AP (63 cm) y hasta el final de la zona Rhizophora mangle aumenta de manera gradual alcanzando valores de 34%. Los elementos de selva muestran una disminución significativa entre el periodo de 1600 a 600 cal AP (74–34 cm) en el cual la selva alcanza porcentajes del 30%. Fabaceae y Ficus sp. son los taxones dominantes entre los elementos de selva constituyendo el 8 y 16% de la suma total de polen de selva. Ficus sp. y Moraceae presentan valores bajos al inicio de este periodo y se recuperan a partir de 1300 cal AP (70 cm) alcanzando valores de hasta 12 y 10% respectivamente. Euphorbiaceae, Rubiaceae y Combretaceae se mantienen a lo largo de este periodo con abundancias de 1–8%; en menor proporción Haematoxylum campechianum, Bignoniaceae y Myrtaceae ≤1%. Amaranthaceae es el elemento con mayores porcentajes, seguido por Poaceae y Asteraceae. Se registra la presencian de Zea mays casi de manera continua desde la base hasta 1300 cal AP (71 cm).
El sedimento en esta zona muestra características geoquímicas variables. Los porcentajes de los elementos de Ti (0.05–0.07%), Al (0.18–0.31%), Fe (0.13–0.20%) y K (0.17–0.27) mostraron una escasa variabilidad, similar a la de la subzona Ib. No obstante, esta estabilidad se vio interrumpida alrededor de 600 cal AP (46 cm) donde los porcentajes de Al, Fe y K muestran un ligero incremento en sus valores. Los valores de Ca (32.16–41.91%) estuvieron por encima del promedio hasta alcanzar los valores más altos de todo el sondeo entre 850–900 cal AP (57–59 cm). La proporción de Ca/Σ (Ti, Fe, Al) muestra incrementos, registrando los dos valores más altos de todo el sondeo, el primero al inicio de la zona (1700 cal AP; 80 cm) alcanzando valores de 123 y el segundo al final de la zona alrededor de 900 cal AP (59 cm) alcanzando valores de 107. No obstante, esta tendencia es brevemente interrumpida alrededor de 1000 cal AP (62 cm) y 600 cal AP (40 cm) con valores de 70.83 y 59.70 respectivamente.
Zona III (41–0 cm; 550 cal AP–Presente)
Conocarpus erectus presentó valores que oscilan entre 1–5%. Sin embrago, en la parte superior de la zona, Avicennia germinans se vuelve dominante. Los elementos de selva presentaron los valores más altos de todo el sondeo (67%). La familia Fabaceae aumenta sus valores >10% hasta alcanzar picos >30% alrededor de 90 cal AP (17 cm). Ficus sp. y Moraceae son relativamente constantes a lo largo de la zona con valores entre 4–23% alcanzando los valores más altos en la superficie del núcleo de sedimento. Las familias Euphorbiaceae y Rubiaceae también se registran a lo largo de este periodo. Brosimum alicastrum, Sapotaceae y Haematoxylum campechianum aparecen de manera constante con valores ≤3%. Polen de la familia Poaceae es el elemento de disturbio con mayor porcentaje (≤17%), seguido por Amaranthaceae y Asteraceae. Se registra la presencia de Zea mays al final de la zona junto con Pinus sp. Croton sp. y Alnus sp. que aparecen frecuentemente a lo largo de este periodo con valores ≤1%.
Al inicio de la zona los porcentajes elementales de Ti (0.05–0.16%), Al (0.18–0.90%), Fe (0.14–0.42%) y K (0.19–0.62) muestran baja variabilidad. Mientras que Ca registra porcentajes por arriba del promedio (4.28–33.92%). No obstante, después de 177 cal AP (23 cm) aumentan los porcentajes de Ti, Al, Fe y K mientras que los valores de Ca disminuyen significativamente por debajo del promedio manteniéndose con valores bajos hasta el final del sondeo, excepto en los sedimentos depositados alrededor de 9 cal AP (10cm). La proporción Ca/Σ (Ti, Fe, Al) muestra disminución excepto en los sedimentos depositados alrededor de 9 cal AP (10 cm) en los cuales se registra un aumento elemental significativo.
DISCUSIÓN
Con la integración del análisis de polen fósil y geoquímica fue posible analizar la historia de los cambios en la vegetación y de las condiciones físicas del ambiente durante los últimos 3200 años para la Laguna Cementerio.
~3200–2500 cal AP
Durante este periodo se registró la mayor abundancia de A. germinans en comparación con el resto de los elementos de manglar. La vegetación dominante es selva (60%). Particularmente alrededor de 3000 cal AP se presentan altas proporciones de elementos que corresponden a selva. Durante el periodo de 3200 a 2800 cal AP se registró una capa de materia orgánica con restos vegetales que sugieren la presencia de vegetación bien establecida en la Laguna Cementerio. Los elementos Ti, Al, Fe y K sugieren un aumento en el aporte clástico entre 3200 y 2900 cal AP. La curva de los elementos geoquímicos sugiere tendencias similares a las variaciones de los taxa de selva, lo cual nos sugiere un aumento en la erosión provocado por el arrastre de los sedimentos hacia el cuerpo de agua. De acuerdo con Roy et al. (2018), los minerales clásticos terrígenos son transportados desde el origen hacia el cuerpo de agua por escorrentía y su concentración puede incrementar en el sedimento durante los regímenes de alta precipitación. De modo contrario, la curva de Ca y Ca/Σ (Ti, Fe, Al) exhibe valores bajos sugiriendo que al inicio del registro las condiciones eran húmedas.
El registro (Figura 5) de la Laguna Cementerio sugieren condiciones húmedas durante el periodo entre 3200 a 2800, mientras que los registros para la Península de Yucatán muestran una disminución en la precipitación como lo indican los resultados para Lago Chichancanab (Hodell et al., 1995), Río Lagartos (Aragón-Moreno et al., 2012; Carrillo-Bastos et al. ,2013), Rio Hondo (Aragón-Moreno et al., 2018a; 2018b, Laguna de Bacalar (Aragón-Moreno et al., 2024) y en la Reserva de la Biosfera de Los Petenes (RBLP) (Gutiérrez-Ayala et al., 2012; Roy et al., 2017) que registraron un régimen de menor precipitación durante este mismo periodo. Estas diferencias probablemente correspondan a una desfase temporal entre los sitios.
En la zona de la cuenca del Cariaco (Haug et al. 2001; Haug et al. 2003) se reportan bajos valores de Ti en el sedimento, asociados a una disminución en la precipitación. De acuerdo con Haug et al. (2001), las variaciones en la precipitación del Megalayiano en la región del Caribe pueden ser explicadas por cambios en la posición media de la ITCZ (Zona de Convergencia Intertropical, ITCZ por sus siglas en inglés). La precipitación en el área de estudio está influenciada por la posición de la ITCZ, la cual se posiciona sobre el área de estudio durante el verano, cuando ésta y la celda de alta presión Bermuda-Azore migran hacia el norte (Malaizé et al., 2011; Carrillo-Bastos et al., 2013).
Para el Megalayiano, Curtis et al. (1996) evaluaron por medio de isótopos de oxígeno de Punta Laguna, cambios de condiciones húmedas (3300–1800) a secas (1800–900) y nuevamente a húmedas (900–0 años cal AP). Estos cambios presentan un patrón que coinciden con nuestros datos para la Laguna Cementerio. Esta fase de humedad reportada al inicio del registro concuerda con la fase tipo Niña que reporta Conroy et al. (2008) para el Junco entre 3800 y 2800 cal AP (Figura 6). Numerosos estudios consideran la ITCZ como la principal fuerza causante de los cambios en el clima de la región. Haug et al. (2001) sugieren que el periodo altamente variable entre 3500 y 2600 cal AP estuvo relacionado con el inicio del aumento de la actividad de ENSO.
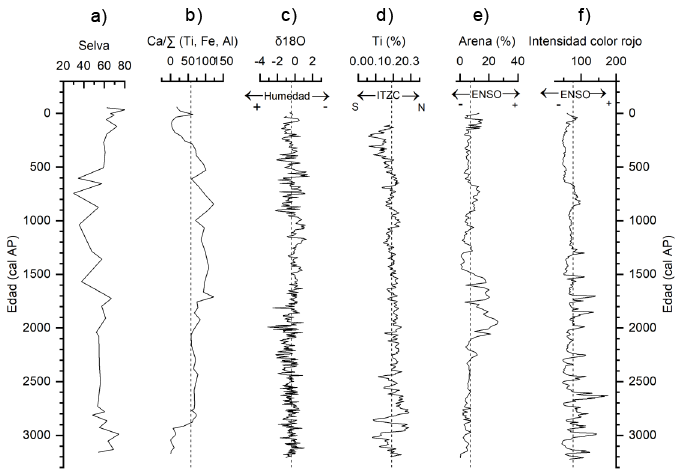
Figura 6. Correlación de los últimos 3200 años comparando (a) porcentaje de vegetación selvática junto con el proxy climático (b) Ca/∑ (Ti, Fe, Al) en la Laguna Cementerio con los registros de (c) Punta Laguna (Curtis et al., 1996), (d) la Cuenca del Cariaco (Haug et al., 2001), (e) Lago el Junco (Conroy et al., 2008) y (f) Laguna Pallcacocha (Moy et al.,2002).
Así mismo, en un registro de isótopos de oxígeno en corales de la isla Navidad en el Pacífico central Woodroffe et al. (2003) observaron que las variaciones interanuales de la temperatura de la superficie del mar y la precipitación durante ENSO fueron menos intensas durante el periodo entre 3800 y 2800 cal AP. Además, durante el Megalayiano, registraron cambios en la magnitud de ENSO que exceden los atribuidos por los cambios solares y que, en lugar de un aumento gradual en las variaciones de ENSO, ha habido al menos un cambio abrupto y no lineal en la amplificación en la variabilidad de ENSO como resultado de una mayor interacción de la convección atmosférica tropical con la Oscilación del Sur.
Algunos autores han registrado el periodo entre 3000 y 2800 cal AP como un periodo altamente variable y anómalo (Woodroffe et al., 2003; Mayewski et al., 2004; Mueller et al., 2009; Malaizé et al., 2011 Aragón-Moreno et al., 2018a). Una alta actividad en los eventos de ENSO y una posición altamente variable de la ITCZ sugieren un efecto combinado de ambos sistemas entre 3800–2800 cal AP (Conroy et al. 2008; Joo-Chang et al. 2015; Aragón-Moreno et al. 2018a). Las diferentes respuestas geográficas durante el periodo entre 3200–2500 cal AP pudieran estar provocadas por variaciones en la dinámica de la corriente de chorro del Caribe (CLLJ, por sus siglas en inglés). La corriente de chorro del Caribe está relacionada con el transporte de grandes cantidades de humedad del Atlántico tropical al mar Caribe, hacia el Golfo de México a través de Centro América hasta la cuenca del Pacífico. Los efectos sobre el Caribe dependen de dos factores: la fuerza y posición del anticiclón de Bermudas-Azores (North Atlantic Subtropical High, NASH por sus siglas en inglés) y por el calentamiento sobre la porción norte de Sudamérica (Cook & Vizy, 2010). Por otra parte, la actividad ciclónica en el Caribe incrementa la cantidad de precipitación sobre algunas zonas del Caribe. En Belice, McCloskey y Keller (2009) registraron una alta actividad ciclónica entre 4500–2500 cal AP. En las Islas de San Martin en el Caribe, Bertran et al., (2004) registraron alta actividad de huracanes entre 4900–2600 cal AP lo cual podría estar relacionado con que en el área de estudio este periodo sea húmedo en comparación con otros sitios de la Península de Yucatán.
Durante este periodo, ocurre el primer registro del polen de maíz (Zea mays) alrededor de 2800 cal AP, lo que podría interpretarse como el inicio de la actividad humana en la región. En Río Lagartos, Aragón-Moreno et al. (2012) reporta el primer registro del maíz alrededor de 3780 cal AP. En Laguna Tzib, Carrillo-Bastos et al. (2010) reporta la presencia del maíz a partir de 3500 cal AP. En lago Cobá, Leyden et al. (1998) reportan la presencia del maíz alrededor de 2850 cal AP. En Laguna de Bacalar, Aragón-Moreno et al. (2024) registra la presencia de actividades agrícolas alrededor de 3500 cal AP, mientras que, para el largo Petén Itzá, Mueller et al. (2009) registraron la presencia del maíz alrededor de 3000 cal AP.
Entre 2800-2500 los taxa de selva indicadores de condiciones húmedas disminuyen mientras que los elementos de disturbio junto con la presencia del maíz incrementa, como lo reportado por Islebe et al., (2022) para algunos sitios de la Península de Yucatán. La dominancia de C. erectus y la disminución de R. mangle son evidencia de la dinámica en la línea de costa, proporcionando evidencias de baja precipitación y disminución en el nivel del mar (Aragón-Moreno et al. 2012). La reducción en el aporte de Ti, Al, Fe y K y una tendencia hacia el aumento en los valores de Ca y Ca/Σ (Ti, Fe, Al) sugieren condiciones de menor precipitación con respecto al inicio de este periodo. De acuerdo con Xiao et al., (2006) valores bajos de calcita están asociados a disminución en la precipitación.
Este periodo de tendencias de diminución en la precipitación se encuentra dentro de los periodos, de acuerdo con Mayewski et al. (2004), que corresponden a cambios climáticos rápidos y significativos y dentro del periodo donde se registró la migración de la ITCZ hacia el sur provocado por una disminución en la actividad solar (Haug et al., 2001). Los datos concuerdan con otros trabajos registrados en la Península de Yucatán, como Punta Laguna (Curtis et al., 1996), Lago Chichancanab (Hodell et al., 2005) y Río Lagartos (Aragón-Moreno et al., 2012). De acuerdo con Aragón-Moreno et al., (2012) durante el periodo entre 3400–2200 cal AP registraron una tendencia de sequía para Río Lagartos. De modo similar, Islebe y Sánchez (2002) y Torrescano-Valle (2007) reportaron una disminución en los elementos de selva que pueden ser interpretados como una reducción de la precipitación.
~2500–1800 cal AP
En este periodo continúa la disminución en la precipitación y se registró una ligera disminución en los elementos de selva en comparación con el periodo anterior (de 55 a 51%). No obstante, se registró la presencia, de Bursera simaruba, Haematoxylum campechianum, Metopium brownei y Buxaceae, aunque en porcentajes bajos. Estos elementos indican periodos de sequía de menor grado. Además, se registra un aumento significativo de Ficus sp. y de acuerdo con Islebe y Sánchez (2002) su presencia puede ser abundante durante un disturbio, por su comportamiento oportunista y tolerante a las variaciones ambientales. Por otra parte, los elementos de manglar se reducen ligeramente, C. erectus muestra dominancia en relación a las demás especies de manglar y apunta a condiciones con tendencias más secas. La presencia de los foraminíferos se limita a la primera zona del registro (entre el 3190 al 2000 cal AP), esta tendencia es similar a lo reportado por Aragón-Moreno et al., (2024) en Laguna Bacalar. La presencia de los foraminíferos es indicativa de cambios en la intrusión marina y niveles intermareales (Joo-Chang et al., 2015). Los elementos asociados a campos abiertos como Poaceae, Amaranthaceae, Convolvulaceae y Zea mays muestran un aumento. Entre 2100 y 1800 cal AP, la curva de carbonato de Ca/Σ (Ti, Fe, Al) presenta un periodo alterno con una ligera disminución para nuevamente incrementar al final del periodo. Cuando se registra una disminución en los valores de Ca y Ca/∑ (Ti, Fe, Al) se detecta la presencia del polen de maíz. De modo similar los valores de calcio y Ca/Σ (Ti, Fe, Al) disminuyen y los valores de Ti, Al, Fe, y K aumentan levemente.
Este intervalo es contemporáneo con las condiciones áridas observadas en la Reserva de la Biosfera de los Petenes entre 2200–1500 cal AP en el cual se registró una disminución considerable en los elementos de selva y de K/Ca (Gutiérrez-Ayala et al., 2012; Roy et al. 2017). En el Río Hondo, Aragón-Moreno et al. (2018a) registraron condiciones secas durante el periodo entre 2600–2100 cal AP en el cual observaron una disminución en Ca/Fe y un incremento significativo de Asteraceae. En la región del Circum-Caribe, la tendencia al incremento de condiciones de sequía, concuerda con lo observado en Haití en el lago Miragoane en donde registraron un periodo de sequías entre 2400 y 1500 cal AP (Hodell et al., 1991). Este periodo de sequía corresponde con un evento de ENSO reportado tanto en Laguna Pallcacocha (Moy et al., 2002) como en Laguna El Junco (Conroy et al., 2008) y una ITCZ estable (Figura 6, Haug et al., 2001).
~1800–550 cal AP
El inicio de esta fase está marcado por la reducción en los taxa de selva y el registro continuo del polen de maíz. Durante este periodo se observaron los porcentajes más bajos de los elementos de selva (42%) como una evidencia de una vegetación más abierta. Se registró un ligero incremento en los elementos de manglar y particularmente C. erectus y A. germinans. Estudios de lluvia de polen y paleoecológicos (Islebe & Sánchez, 2002; Escarraga-Paredes et al., 2014) sugieren que C. erectus es una especie que aparece en condiciones de menor precipitación; además la presencia de Avicennia germinans sugiere un ambiente salino o hipersalino causado por cambios intermareales. Los elementos de disturbio como Poaceae, Amaranthaceae, Asteraceae y Convolvulaceae se mantienen a lo largo de este periodo respaldando la interpretación de condiciones progresivamente más secas.
Alrededor de 1700 cal AP durante el Clásico temprano (250 d. C) la curva de Ca/Σ (Ti, Fe, Al) sugiere un evento de sequía severa, seguido de fases alternadas de periodos secos y húmedos. Esta sequía de corta duración (~50 años), pueden explicar la reaparición de maíz (Islebe et al., 2022) y posterior recuperación de la selva. Los cultivos aparecen cuando se registran condiciones de menor precipitación de manera casi continua a lo largo de este periodo. Islebe et al., (2022) hipotetizan que algunas regiones de la Península de Yucatán la presencia del polen de maíz se detecta en condiciones climáticas menos favorables. Los autores sugieren que el maíz era un recurso alimenticio utilizado durante los periodos de sequía y hambruna, cuando otros cultivos escaseaban. Las condiciones de sequía regresan alrededor de 850 cal AP (1100 d.C.) en el cual se registraron los porcentajes más bajos de selva de todo el sondeo. En este periodo el conjunto de taxa muestran que la vegetación era un mosaico compuesto por selva de carácter secundario y abierto (Ficus sp. y Fabaceae), con vegetación de disturbio (Amaranthaceae, Poaceae y Convolvulaceae) y cultivos (Z. mays).
Las condiciones secas durante este periodo se han identificado en otros registros climáticos en la Península de Yucatán y la región del Caribe. Carrillo-Bastos et al. (2010) reportan para el lago Tzib condiciones de menor precipitación después de 1900 cal AP, registrando un pico de sequía alrededor de 1200 cal AP. Estas condiciones de sequía persistieron hasta 850 cal AP. Los autores mencionan que a pesar de que el inicio del Megalayiano es una fase húmeda, se distingue una fase seca durante el periodo Clásico (250–900 d.C). Por otro lado, Islebe y Sánchez (2002), en Puerto Morelos, registran un aumento de C. erectus entre 1500 cal AP y 1250 cal AP como un reflejo de condiciones de sequía durante ese periodo. Hodell et al. (2005), en el lago Chichancanab registraron dos sequías durante el periodo Clásico terminal, la segunda sequía inicio en 920 d.C (1030 cal AP) y culminó alrededor de 1100 d.C (850 cal AP). Dentro de estas fases de sequías, el clima estaba marcado por periodos alternos de condiciones secas y húmedas con un intervalo recurrente en las sequías de ~50 años. Curtis et al., (1996) reportan dos periodos de sequía de 750 a 850 d.C. (1200–1400 cal AP) y de 910 a 990 d.C. (1040–960 cal AP) en Punta Laguna con un periodo húmedo entre 850 y 910 d.C (1100–1040 cal AP) Además, proponen que el colapso de la cultura maya ocurrió entre 800–1000 d. C (1150–950 cal AP) Para este mismo sito, Hodell et al., (2007) observaron que el clima fue más seco en el periodo Clásico que en el Preclásico. En el registro de la cuenca del Cariaco no se registró variabilidad en los valores de Ti durante el periodo entre 2500–1000 cal AP (Haug et al., 2001) cuando los registros de variabilidad de ENSO (Moy et al., 2002, Conroy et al., 2008) muestran mayor frecuencia, magnitud y duración en los eventos de ENSO lo que sugiere una disociación entre la ITCZ y ENSO. Por lo que, diferentes autores han explorado otros mecanismos que permitan explicar esta variabilidad climática.
Woodroffe et al. (2003) sugieren que las variaciones interanuales de la temperatura de la superficie del mar y la precipitación asociadas a ENSO fueron menos intensas entre 3800 y 2800 cal AP, pero más pronunciadas alrededor de 1700 cal AP. Los registros de laguna Pallcacocha y Laguna El Junco sugieren que entre 2000 y 1000 cal AP ocurrió un periodo de frecuencia extremadamente alta de eventos de ENSO y tal vez la más alta de todo el Holoceno (Moy et al., 2002; Conroy et al., 2008). Por otra parte, Wu et al. (2017) mencionan que estos periodos de menor precipitación podrían estar asociados a un cambio pronunciado en el gradiente de la temperatura superficial del mar y la salinidad afectado la corriente de chorro del Caribe (CLLJ, por sus siglas en inglés) con consecuencias en la cantidad de precipitación en Mesoamérica. Este cambio pronunciado en el gradiente de la temperatura superficial de mar, también podría estar relacionado con cambios en la circulación de Retorno Meridional de Atlántico (AMOC, por sus siglas en inglés). La corriente de chorro del Caribe es uno de los contribuyentes de sequías y anomalías en la precipitación debido a los cambios en la divergencia del flujo de humedad en el Caribe. Los autores sugieren que la ITCZ no puede ser el único mecanismo para explicar las anomalías hidrológicas observadas en la Península de Yucatán especialmente durante el periodo Clásico terminal y en su lugar, se sugiere que estas anomalías negativas en la precipitación en Mesoamérica durante este periodo pudieron haber tenido la influencia de eventos cálidos de ENSO, los cuales están vinculados con la corriente de chorro del Caribe.
Posteriormente a esta fase de sequía, alrededor de 700 cal AP la curva de Ca/Σ (Ti, Fe, Al) muestra un incremento en la humedad y se registra un aumento significativo en los elementos de selva hasta el final del periodo. Para la Península de Yucatán, Carrillo-Bastos et al. (2010) registraron recuperación de los elementos de selva a partir de 750 cal AP. Aragón-Moreno et al. (2012) reportan un aumento en Moraceae y Ficus sp. a partir de 770 cal AP, lo que sugiere un incremento en los porcentajes de selva como respuesta al aumento de las condiciones húmedas. Para la Reserva de la Biosfera de Los Petenes, Roy et al. (2017) detectan un aumento en K/Ca, así como un aumento en la vegetación como una respuesta a un periodo de incremento de la humedad entre 825–700 cal AP.
~550 cal AP–Presente
Desde el inicio de este periodo hasta alrededor de 33 cal AP se presentan condiciones favorables para el aumento de los elementos de selva y vegetación de manglar (17%), así como reducción en los elementos de disturbio (20%). En las curvas de polen la vegetación selvática muestra recuperación y alcanzan los valores máximos (63%) de todo el sondeo. Se registran altos porcentajes de Ficus sp., Moraceae, Fabaceae y Euphorbiaceae, y en menores proporciones Bursera simaruba, Rubiaceae, Haematoxylum campechianum, Myrtaceae y Thrinax radiata. Esta diversidad de taxa y la proporción en que se presentan indican el dominio de selva con mayor madurez (Torrescano-Valle, 2007; Carrillo-Bastos, 2008; Escarraga-Paredes et al., 2014). Las especies de manglar se recuperan de manera progresiva a lo largo de este periodo, las especies dominantes son Avicennia germinans y Rhizophora mangle (3–16%). Las condiciones de precipitación abundante se reflejan en las curvas de los elementos Ti, Al, Fe y K con una tendencia de aumento, lo que sugiere que el incremento en la precipitación ocasionó el aporte de sedimentos a la laguna. Mientras que la curva de Ca/Σ (Ti, Fe, Al) muestra una disminución en sus valores, apoyando la idea de condiciones húmedas.
Este periodo de condiciones favorables es consistente con lo reportado por Aragón-Moreno et al. (2012) donde observaron un aumento en los elementos de selva indicando el regreso de las condiciones de humedad en la región. Carrillo-Bastos et al. (2010) reportaron un aumento en la precipitación alrededor de 750 cal AP al centro de la Península de Yucatán seguido de la expansión de la selva. En la región de Peten campechano Torrescano-Valle et al. (2023) reportaron el aumento en las concentraciones elementales, lo que sugiere el regreso de las condiciones húmedas entre 1350 y 1850 d.C. durante el posclásico (600–100 cal AP). En lago Chichancanab el regreso de las condiciones húmedas se registró después de 1100 d.C. (850 cal AP) (Hodell et al., 2005).
CONCLUSIONES
Mediante el análisis del polen fósil y geoquímica obtenido de los sedimentos, se determinó la historia de la vegetación y las condiciones del medio físico en las cercanías de la Laguna Cementerio, Xcalak Quintana Roo durante los últimos 3200 años. La proporción de Ca/Σ (Ti, Fe, Al) se utilizó para reconstruir variaciones en la precipitación. Durante el inicio del sondeo se registraron condiciones húmedas, en donde la vegetación dominante era la selva compuesta principalmente por elementos de Moraceae, Fabaceae, Euphorbiaceae, Rubiaceae y Bursera simaruba. Así, los cambios registrados en la vegetación durante los últimos 3200 años cal AP muestran una respuesta a la dinámica hidrológica de la zona. Las distintas respuestas climáticas en los distintos sitios de la Península de Yucatán durante este periodo podrían estar relacionadas con variaciones en la dinámica de la corriente de chorro del Caribe y la actividad ciclónica.
Se lograron identificar dos sequías intensas, la primera alrededor de 1700 cal AP y la segunda alrededor de 850 cal AP, esta última podría coincidir con las sequías relacionadas con el colapso de la Cultura Maya. Durante el periodo de sequía intensa alrededor de 1700 cal AP la vegetación arbórea se redujo y adquirió un carácter más abierto. Los resultados sugieren que el clima y la vegetación podrían estar fuertemente influenciados por cambios en la posición de la ITCZ, actividad de ENSO, cambios en el gradiente de temperaturas superficiales y la salinidad que tiene un efecto sobre la corriente de chorro del Caribe.
El registro de Zea mays permitió identificar la presencia de actividades agrícolas cerca de la zona en el Preclásico medio (alrededor de 2800 cal AP) y que continuaron durante del Clásico hasta el Posclásico tardío cuando se registró una disminución en la precipitación. En este periodo se observó la presión simultánea de ambos factores sobre la cobertura vegetal, la sequía y la agricultura, dando como resultado una transformación del paisaje (carácter abierto).
MATERIAL SUPLEMENTARIO
La Tabla Suplementaria S1 donde se muestra la concentración de elementos geoquímicos y proxy climatico en Laguna Cementerio, se puede descargar en el portal web de la Revista <rmcg.unam.mx>, dentro de la página de vista previa de este articulo <http://dx.doi.org/10.22201/igc.20072902e.2025.1.1854>
Agradecimientos. Agradecemos la revisión y comentarios por Carmen Isela Ortega Rosas y un revisor anónimo.
Contribución de los autores. C.B.U.R. redacción, revisión, ediciones del manuscrito, borrador original, visualización, investigación, metodología, software, análisis formal. A.C.B. redacción, revisión, ediciones del manuscrito, borrador original, visualización, conceptualización, metodología, software, análisis formal, recursos, administración del proyecto, supervisión, validación. C.G.S. revisión, ediciones del manuscrito. A.A.A.M. revisión, ediciones del manuscrito. G.I. revisión, ediciones del manuscrito.
Financiamiento. Este proyecto fue financiado por el Tecnológico Nacional de México a través del proyecto “Dinámica de incendios y variabilidad climática durante los últimos 2000 años en el sur de Quintana Roo” (clave: 17853.23-P).
Política de disponibilidad de datos. Datos disponibles previa solicitud justificada a los autores.
Declaración de conflicto de intereses. Los autores declaran que no tienen conflictos de intereses.
REFERENCIAS
Álvarez, M. (1954). Exploración geológica preliminar del rio Hondo, Quintana Roo, México. Boletín de la Asociación Mexicana de Geólogos Petroleros, 5(5-6), 207–213
Aragón-Moreno, A. A., Islebe, G. A., & Torrescano-Valle, N. (2012). A ˜3800-Yr, high resolution record of vegetation and climate change on the North Coast of the Yucatan Peninsula. Review of Paleobotany and Palynology, 178, 35–42. https://doi.org/10.1016/j.revpalbo.2012.04.002
Aragón-Moreno, A. A., Islebe, G. A., Roy, P. D., Torrescano-Valle, N., & Mueller, A. D., (2018a). Climate forcings on vegetation of the southeastern Yucatán Peninsula (Mexico) during the middle to late Holocene: Palaeogeography, Palaeoclimatology, Palaeoecology, 495, 214–226. https://doi.org/10.1016/j.palaeo.2018.01.014
Aragón-Moreno, A., Islebe, G., Torrescano-Valle, N., & Arellano-Verdejo, J., (2018b) Middle and late Holocene mangrove dynamics of the Yucatan Peninsula, Mexico. Journal of South American Earth Sciences, 85, 307–311. https://doi.org/10.1016/j.jsames.2018.05.015
Aragón-Moreno, A. A., Islebe, G. A., Torrescano-Valle, N., Hernandez-Arana, H., Carrillo, A., & Friedel, R. (2024). Mangrove dynamics driven by Late Holocene drought at Laguna Bacalar, Mexico. Quaternary Environments and Human, 2, 100023. https://doi.org/10.1016/j.qeh.2024.100023
Bautista, F., & Palacio, G. (2011). Parte III. Regionalización edáfica del territorio de México. Capítulo 24. Península de Yucatán. En P., Krasilnikov, F.J., Jiménez, T., Reyna, & N.E., García, (eds), Geografía de Suelos de México (355–406 pp). Universidad Nacional Autónoma de México.
Bautista, F., Palacio-Aponte, G., Quintana, P., & Zinck, J.A. (2011). Spatial distribution and development of soils in tropical karst areas from the Peninsula of Yucatan, Mexico. Geomorphology, 135(3-4), 308–321. https://doi.org/10.1016/j.geomorph.2011.02.014
Bertran, P., Bonnissent, D., Imbert, D., Lozouet, P., Serrand, N., & Stouvenot, C. (2004). Paleoclimat des Petites Antilles depuis 4000 BP:l’enregistrement de la lagune de Grand-Case a Saint-Martin. C.R. Geoscience, 336, 1501–1510. https://doi.org/10.1016/j.crte.2004.09.009
Birks, H. J. B. (2019). Contributions of Quaternary botany to modern ecology and biogeography: Plant Ecology and Diversity, 12(3-4), 189–385. https://doi.org/10.1080/17550874.2019.1646831
Blaauw, M., & Christen, J.A. (2011). Flexible paleoclimatic age-depth models using autoregressive gamma process. Bayesian Analysis, 6(3), 457–474. https://doi.org/ 10.1214/ba/1339616472
Brewer, S, Guiot J., & Barboni D. (2013). Use of Pollen as Climate Proxies, in A. E., Scott Mock C. J. (eds), Encyclopedia of Quaternary Science. (pp. 805-815)Elsevier, Oxford. https://doi.org/10.1016/B978-0-444-53643-3.00180-1
Can-Canales, C., Islebe, G., Carrillo, A., Torrescano-Valle, N., & Aragón-Moreno, A. (2023). Fungal spores in Caribbean mangrove sediments, dataset from southeastern Mexico. Data in Brief, 51, 109631. https://doi.org/10.1016/j.dib.2023.109631
Carrillo-Bastos, A. (2008). Variación climática y ecológica durante el Holoceno Medio-Tardío en la Península de Yucatán, un enfoque geo-espacial. [Tesis de Maestría] El Colegio de la Frontera Sur.
Carrillo-Bastos, A., Islebe, G. A., Torrescano-Valle, N., & González, N. E., (2010). Holocene vegetation and climate history of central Quintana Roo, Yucatán Península, Mexico. Review of Palaeobotany and Palynology, 160, 189–196. https://doi.org/10.1016/j.revpalbo.2010.02.013.
Carrillo-Bastos, A., Islebe, G.A., & Torrescano-Valle N. (2013). 3800 Years of Quantitative Precipitation Reconstruction from the Northwest Yucatan Peninsula. PLoS ONE, 8(12). https://doi.org/10.1371/journal.pone.0084333
Chevalier M., Basil A.S., Davis, O. H., Heikki, S, Brian, M., Chase, K. G., Terri, L., Richard, J. T., Finsinger, W., Guiot, J., Kühl, N., Maezumi S. Y., Tipton, J.R., Carter, V. A., Brussel, T., Phelps L.N., Dawson, A., Zanon, M., Vallé, F., Nolan, C., Mauri, A., de Vernal, A., Izumi, K., Holmström, L., Marsicek, J., Goring, S., Sommer, P.S., Chaput M., & Kupriyanov, D., (2020). Pollen-based climate reconstruction techniques for late Quaternary studies. Earth-Science Reviews, 210, 103384. https://doi.org/10.1016/j.earscirev.2020.103384
Cook, K., & Vizy, E. (2010). Hydrodynamics of the Caribbean Low-Level Jet and Its Relationship to Precipitation. Journal of Climate, 23, 1477–1494. https://doi.org/10.1175/2009JCLI3210.1
Comisión Nacional de Áreas Naturales Protegidas (CONANP) (2004). Programa de Manejo Parque Nacional Arrecifes de Xcalak, México. Disponible en http://www.conanp.gob.mx/que_hacemos/pdf/programas_manejo/Xcalak_ok.pdf, (consultado: 10/11/2024).
Conroy, J. L., Overpeck, J. T., Cole, J. E., Shanahan, T. M., & Kannan M. S. (2008). Holocene changes in eastern tropical Pacific climate inferred from a Galapagos Lake sediment record. Quaternary Science Reviews, 27(11-12), 1166–1180. https://doi.org/10.1016/j.quascirev.2008.02.015
Curtis, J. H., Hodell, D. A., & Brenner, M. (1996). Climate variability on the Yucatan Peninsula (Mexico) during the past 3500 yr, and implications for Maya cultural evolution. Quaternary Research, 46(1), 37–47. https://doi.org/10.1006/qres.1996.0042
Ellison, J. C. (2008). Long-term retrospection on mangrove development using sediment cores and pollen analysis: A review. Aquatic Botany, 89(2), 93–104. https://doi.org/10.1016/j.aquabot.2008.02.007
Escarraga-Paredes, D., Torrescano-Valle, N., & Islebe, G. (2014). Análisis de la relación vegetación-lluvia de polen actual de las comunidades vegetales en el noroeste de la península de Yucatán, México. Polibotánica, 38, 27–52.
Faegri, K., & Iversen, J. (1989). Textbook of pollen analysis (4th ed.). En K., Faegri, P.E., Kaland, K., Krzywinski (Eds.). Chichester, John Wiley, 354 pp.
Fragoso-Servón, P., Bautista, F., Fraustro, O., & Pereira A. (2014). Características de las depresiones kárstica (forma, tamaño y densidad) a escala 1: 50,000 y sus tipos de inundación en el estado de Quintana Roo, México. Revista Mexicana de Ciencias Geológicas, 31(1), 127–137.
Friedel, T., Johanson, E. N., & Horn, S. P. (2024). Post-Conquest vegetation and fire dynamics at Laguna Carse, Costa Rica: A pollen and microscopic charcoal record. Vegetation History and Archaeobotany, 33, 595–606. https://doi.org/10.1007/s00334-023-00981-7
García, E. (1973). Modificación al sistema de clasificación climática de Koppen (Segunda Edición). Comisión Nacional para el estudio de la Biodiversidad (CONABIO), 97 pp.
Grimm, E. C. (1987). CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method on incremental sum of square. Computer & Geoscience, 13(1), 13–35. https://doi.org/10.1016/0098-3004(87)90022-7
Grimm, E. C. (2016). Tilia (v3.0.3) [software]. Illinois State Museum. Research and Collection Center, Springfield. Disponible en https://www.neotomadb.org/apps/tilia
Gutiérrez-Ayala, L. V., Torrescano-Valle, N., & Islebe, G. A. (2012). Reconstrucción paleoambiental del Holoceno Tardío de la reserva los Petenes, Península de Yucatán, México. Revista Mexicana de Ciencias Geológicas, 29(3), 749–763.
Haug, G. H., Hughen, K. A., Sigman, D. M., Peterson, L. C., & Rohl, U. (2001). Southward migration of the Intertropical Convergence Zone through the Holocene. Science, 293, 1304–1308. https://doi.org/10.1126/science.1059725
Haug, G. H., Gunther, D., Peterson, L.C., Sigman, D. M., Hughen, K. A., & Aeschlimann B. (2003). Climate and the collapse of Maya civilization. Science, 299, 1731–1735. https://doi.org/10.1126/science.1080444
Hirales-Cota, M., Espinoza-Avalos, J., Schmook, B., Ruiz-Luna, A., & Ramos-Reyes, R., (2010). Drivers of mangrove deforestation in Mahahual-Xcalak, Quintana Roo, Southeastern Mexico. Ciencias Marinas, 36(2), 147–159. https://doi.org/10.7773/cm.v36i2.165
Hodell, D., Curtis, J., Higuera-Gundy, A., Brenner, M., Jones, G., Binford, M., & Dorsey, K. T. (1991). Reconstruction of Caribbean Climate Change over the Past 10,500 Years. Nature, 352, 790–793. https://doi.org/10.1038/352790a0
Hodell, D. A., Curtis, J. H., & Brenner, M. (1995). Possible role of climate in the collapse of Classic Maya civilization. Nature, 375, 391–394. https://doi.org/10.1038/375391a0
Hodell, D. A., Brenner, M., Curtis, J. H., & Guilderson, T. (2001). Solar forcing of drought frequency in the Maya lowlands. Science, 292(5520), 1367–1370. https://doi.org/10.1126/science.1057759
Hodell, D. A., Brenner, M., & Curtis, J. H. (2005). Terminal Classic drought in the northern Maya Lowlands inferred from multiple sediment cores in Lake Chichancanab (Mexico). Quaternary Science Reviews, 24(12-13), 1413–1427. https://doi.org/10.1016/j.quascirev.2004.10.013
Hodell, D., Brenner, M., & Curtis, J. (2007). Climate and cultural history of the Northeastern Yucatan Peninsula, Quintana Roo, Mexico. Climatic Change, 83, 215–240. https://doi.org/10.1007/s10584-006-9177-4
Islebe, G., & Sánchez, O. (2002). History of late Holocene vegetation at Quintana Roo, Caribbean coast of Mexico. Plant Ecology, 160, 187–192. https://doi.org/10.1023/A:1015865932012
Islebe, G. A., Sánchez, O., Váldez-Hernandez, M., & Weissenberger, H. (2015). Distribution of vegetation types. En G. A., Islebe, S., Calmé, J. L. J., León-Cortés, B. Schmook, (eds.). Biodiversity and conservation of the Yucatán Peninsula (39–53 pp). Springer International Publishing.
Islebe, G. A., Torrescano-Valle, N., Valdez-Hernández, M., Carrillo-Bastos, A., & Aragón-Moreno, A. A. (2022). Maize and ancient Maya droughts. Scientific Reports, 12(22272), https://doi.org/10.1038/s41598-022-26761-3
Joo-Chang, J. C., Islebe, G. A., Torrescano-Valle, N. (2015). Mangrove history during middle-and late- Holocene in Pacific south-eastern Mexico. Holocene, 25(4), 651–662. https://doi.org/10.1177/0959683614566217
Leyden, B. W., Brenner, M., & Dahlin, B. H., (1998). Cultural and climatic history of Cobá, a lowland Maya city in Quintana Roo, Mexico. Quaternary Research, 49(1), 111–122. https://doi.org/10.1006/qres.1997.1941
Liu, F., Ding, J., Zeng, J., Wang, C., Wu, B., Yan, Q., He, Z., Shu, L. (2024). Mangrove sediments are environmental hotspots for pathogenic protists. Journal of Hazardous Materials, 467(133643). https://doi.org/10.1016/j.jhazmat.2024.133643
Lozano-García, Ma. Del S., & Martínez-Hernández, E. (1990). Palinología de los Tuxtlas: Especies arbóreas. Instituto de Biología, Universidad Nacional Autonoma de México, 120 pp.
Lovelock, C. E., Cahoon, D. R., Friess, D. A., Guntenspergen, G. R., Krauss, K. W., Reef, R., Rogers, K., Saunders, M. L., Sidik, F., Swales, A., Saintilan, N., Thuyen, L. X., Triet, T, (2015). The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature, 526, 559–563. https://doi.org/10.1038/nature15538
McCloskey, T. A., Keller, G. (2009). 5000-year sedimentary record of hurricane strikes on the central coast of Belize. Quaternary International, 195(1-2), 53–68. https://doi.org/10.1016/j.quaint.2008.03.003
Malaizé, B., Bertran, P., Carbone, P., Bonnissent, D., Charlier, K., Galop, D., Imbert, D., Serrand, N., Stouvenot, Ch., & Pujol, C. (2011). Hurricanes and climate in the Caribbean during the past 3700 years BP B. The Holocene, 21(6), 911–924. https://doi.org/10.1177/0959683611400198
Mayewski, P. A., Rohling, E. E., Curt Stager, J., Karlén, W., Maasch, K. A., David Meeker, L., Meyerson, E. A., Gasse, F., van Kreveld, S., Holmgren, K., Lee-Thorp, J., Rosqvist, G., Rack, F., Staubwasser, M., Schneider, R. R., Steig, E. J. (2004). Holocene climate variability. Quaternary Research, 62(3), 243–255. https://doi.org/10.1016/j.yqres.2004.07.001
Monacci, N. M., Meier-Grünhagen, U., Finney, B. P., Behling, H., Wooller, M. J. (2009). Mangrove ecosystem changes during the Holocene at Spanish Lookout Cay, Belize. Palaeogeography, Palaeoclimatology, and Palaeoecology, 280(1-2), 37–46. https://doi.org/10.1016/j.palaeo.2009.05.013
Monacci, N.M., Meier-Grünhagen, U., Finney, B.P., Behling, H., & Wooller, M.J. (2011). Laleoecology of mangroves along the Sibun River, Belize.Quaternary Research, 76(2), 220–228. https://doi.org/10.1016/j.yqres.2011.06.001
Moy, C.M., Seltzer, G.O., Rodbell, D.T., Anderson, D.M. (2002). Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature, 420, 162–165. https://doi.org/10.1038/nature01194
Mueller, A. D., Islebe, G. A., Hillesheim, M. B., Grzesik, D. A., Anselmetti, F. S., Ariztegui, D., Brenner, M., Curtis, J. H., Hodell, D. A., & Venz, K. A. (2009). Climate drying and associated forest decline in the lowlands of northern Guatemala during the late Holocene. Quaternary Research, 71(2), 133–141. https://doi.org/10.1016/j.yqres.2008.10.002
Ordoñez-Crespo, L., & García-Rodríguez, M. (2010). Formas kársticas comunes de los cenotes del estado de Quintana Roo (México). M+A Revista electrónica de Medio Ambiente, 9, 15–35.
Palacios-Chávez, R., Ludlow-Wiechers, B., & Villanueva-Gutiérrez, R. (1991). Flora palinológica de la Reserva de la Biosfera de Sian Ka ́an, Quintana Roo, México. Centro de Investigaciones de Quintana Roo.
Phuphumirat W., Zetter R., Hofmann,C. C., Ferguson, D. K. (2015). Pollen degradation in mangrove sediments: A short-term experiment. Review of Palaeobotany and Palynology, 221, 106–116. https://doi.org/10.1016/j.revpalbo.2015.06.004
Punwong, P., Sritrairat, S., Selby, K., Marchant, R., Pumijumnong, N., & Traiperm, P. (2018). An 800-year record of mangrove dynamics and human activities in the upper Gulf of Thailand. Vegetation History and Archaeobotany, 27, 535–549. https://doi.org/10.1007/s00334-017-0651-x
Quiroz-Jiménez, J. D., & Roy, P. D. (2017). Evaluation of geochemical data by two different XRF spectometers in sediments from the Santiaguillo Basin (State of Durango, Mexico). Geofísica Internacional, 56(3), 305–315. https://doi.org/10.22201/igeof.00167169p.2017.56.3.1819
R Core Team (2023). R: A language and environment for statistical computing [software]. R Foundation for statistical computing, Vienna, Austria, https://www.R-project.org
Reimer, P. J., Augustin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Ramsey, C. B., Butzin, M., Cheng, H., Edwards, R. L., Friedrich, M., Grootes, P. M., Guilderson, T. P., Hajdas, I., Hearton, T. J. Hogg, A. G., Hughen, K. A., Kromer, B., Manning, S. W., Muscheler, R., Palmer, J. G., Pearson, C., van der Plicht, J., Reimer, R. W., Richards, D. A., Scott, E. M., Southon, J. R., Turney, C. S. M., Wacher, L., Adolphi, F., Buntgen, U., Capano, M., Fahrni, S. M., Fogtmann-Schulz, A., Friedrich, R., Kohler, P., Kudsk, S., Miyake, F., Olsen, J., Reinig, F., Sakamoto, M., Sookdeo, A., & Talamo, S. (2020). The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon, 62(4), 725–757. https://doi.org/10.1017/RDC.2020.41
Roberts, N., 2014. The Holocene. John Wiley and Sons, 378 pp.
Roy, P. D., Torrescano-Valle, N., Islebe, G. A., & Gutiérrez‐Ayala, L. V., (2017). Late Holocene hydroclimate of the western Yucatan Peninsula (Mexico). Journal of Quaternary Science, 32(8), 1112–1120. https://doi.org/10.1002/jqs.2988
Roy, P. D., Torrescano-Valle, N., Escarraga-Paredes, D. S., Vela-Pelaez, A. A., Lozano-Santacruz, R. (2018). Comparison of elemental concentration in near-surface late Holocene sediments and precipitation regimes of the Yucatán Peninsula (Mexico): a preliminary study. Boletín Geológico y Minero, 129(4), 693–706. https://doi.org/10.21701/bolgeomin.129.4.006
Sefton, J. P., & Woodroffe, S. A. (2021) Assessing the use of mangrove pollen as a quantitative sea-level indicator on Mahé, Seychelles. Journal of Quaternary Science, 36(2), 311–323. https://doi.org/10.1002/jqs.3272
Taylor, S. R., & McLennan S. M., (1985). The continental crust: its composition and evolution. Oxford, Blackwell Scientific Publications, 312 pp.
Torrescano-Valle N. (2007). Reconstrucción paleoambiental del Holoceno Medio-Tardío en la parte centro-sur de la Península de Yucatán, México [Tesis de Doctorado]. El Colegio de la Frontera Sur, 94 pp.
Torrescano-Valle, N., & Islebe, G. A. (2006). Tropical Forest and mangrove history of southeastern Mexico: a 5000-yr pollen record and implications for sea level rise. Vegetation History and Archaeobotany, 15, 191–195. https://doi.org/10.1007/s00334-005-0007-9
Torrescano-Valle, N., & Islebe, G. (2012). Mangroves of Southeastern Mexico: Paleoecology and Conservation. The open Geography Journal, 5, 6–15. https://doi.org/10.2174/1874923201205010006
Torrescano-Valle, N., Folan, W., Yanez-Montalvo, A., & Gunn, J. (2023). Climate and agricultural history from the Petén Campechano in the Late Holocene Maya lowlands of southern Mexico. Vegetation History and Archaeobotany, 32, 387–403. https://doi.org/10.1007/s00334-023-00914-4
Urrego, L. E., Bernal, G., & Polanía, J. (2009). Comparison of pollen distribution patterns in surface sediments of a Colombian Caribbean mangrove with geomorphology and vegetation. Review of Palaeobotany and Palynology, 156(3-4), 358–375. https://doi.org/10.1016/j.revpalbo.2009.04.004
Walker, M., Gibbard, P., Head, M. J., Berkelhammer, M., Bjorck, S., Cheng, H., Cwynar L.C., Fisher, D., Gkinis, V., Long, A., Lowe, J., Newnham, R., Rasmussen, S.O., & Weiss H. (2019). Formal Subdivision of the Holocene Series/Epoch: A Summary: Journal of the Geological Society of India, 93, 135–141. https://doi.org/10.1007/s12594-019-1141-9
Woodroffe, C. D., Beech, M. R., & Gagan, M. K., (2003). Mid-late Holocene El Niño variability in the equatorial Pacific from coral microatolls. Geophysical Research Letters, 30(7), 1358. https://doi.org/10.1029/2002GL015868
Wooller, M. J, Morgan, R., Fowell, S., Behling, H., Fogel, M. (2007). A multiproxy peat record of Holocene mangrove paleoecology from Twin Cays, Belize. The Holocene, 17(8), 1129-1139. https://doi.org/10.1177/0959683607082553
Wooller, M. J., Behling, H., Guerrero, J. L., Jantz, N., Zweigert, M. E. (2009). Late Holocene hydrologic and vegetation changes at Turneffe Atoll, Belize, compared with records from mainland Central America and Mexico. Palaios, 24(9-10), 650–665. https://doi.org/10.2110/palo.2009.p09-036r
Wu, H. C., Felis, T., Scholz, D., Giry C., Kölling, M., Jochum K. P., Scheffers S. R. (2017). Changes to Yucatán Peninsula precipitation associated with salinity and temperature extremes of the Caribbean Sea during the Maya civilization collapse. Scientific Reports, 7, (15825). https://doi.org/10.1038/s41598-017-15942-0
Vidal-Zepeda, R., (2005). Las regiones climáticas de México. UNAM, México, Colecciones de Temas Selectos de Geografía de México, temas selectos de Geografía de México, Investigaciones Geográficas. Universidad Nacional de México, Instituto de Geografía, 212 pp.
Xiao J., Wu J., B. Liang W., Nakamura T., Liu B., Inouchi Y. (2006). Holocene Climate changes in the monsoon & Arid transition reflected by carbon concentration in Daihai Lake of Inner Mongolia. The Holocene 16(4), 551–560. https://doi.org/10.1191/0959683606hl950rp
Yan, K., Wang, Cl., Mischke, S., Wang, J. V., Shen, L. J., Yu, X. C, Meng, L. Y. (2021). Major and trace-element geochemistry of Late Cretaceous clastic rocks in the Jitai Basin, southeast China. Scientific Reports, 11, 13846. https://doi.org/10.1038/s41598-021-93125-8
Editores:
Natalia Pardo Villaveces
Luigi Solari
Pilar Navas-Parejo
Copyright
© 2025 The Author(s).
This is an open-access article published and distributed by the Universidad Nacional Autónoma de México under the terms of a Creative Commons Attribution 4.0 International License (CC BY) which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
